 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98(Suppl. I), January 2003, pp. 71-94 Terrestrial Mammal Feces: a Morphometric Summary and Description Marcia Chame
Laboratório de Ecologia, Departamento de Endemias, Escola Nacional de Saúde Pública-Fiocruz, Rua Leopoldo Bulhões 1480, 21240-210 Rio de Janeiro, RJ, Brasil Fax: +55-21-2257.3946. E-mail: mchame@ensp.fiocruz.br Received 8 November 2001 Code Number: oc03014 The study of feces of terrestrial mammals brings out biological and ecological data such as the species presence, diet, behaviour, territory, parasitic fauna, and home-range use, which can be applied for conservation projects and support paleoecological research that use coprolites as the main source of study. Although the new biotechnological techniques allow more accurate data, the diagnosis based on morphometric analyses permits the primary identification of the taxonomic group origin to support the best choice of subsequent analyses. We present the compilation list of fecal shape and measurements available in the literature published in North America, Eastern and Southern Africa, Europe, and new data from Brazil. Shape and diameters are the best characteristics for taxonomic identification. Feces were assembled in 9 groups that reflect the Order, sometimes the Family, and even their common origin.
Key words: feces - coprolites - terrestrial mammals - fauna Tracking is probably the oldest science (Liebenberg 1990). By looking for signs left by animals we learn to observe useful details to hunt them or to avoid them. Tracking reveals the age of marks left by animals and the natural behaviour of animals without the influence of the observer (Wemmer et al. 1996). As a non-invasive method, it constitutes an important tool for studying threatened species or animals difficult to observe and trap. It can be applied as well for studying rare and nocturnal animals. Although it requires observers who are well trained with sharp sensitivity, its low cost and accessible technology turn tracking into a good choice for field studies.
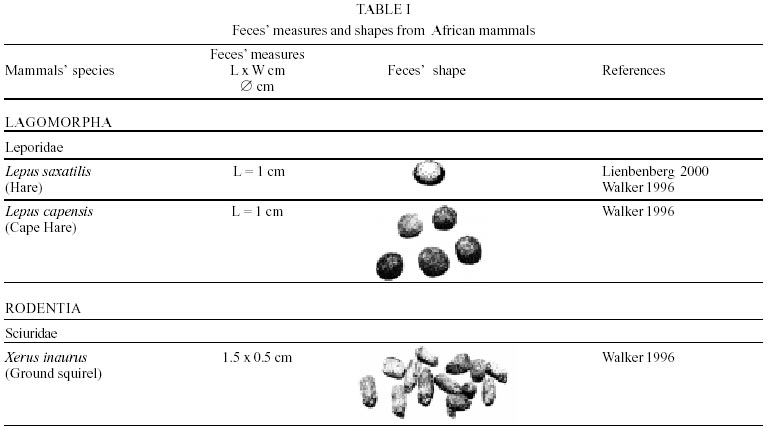
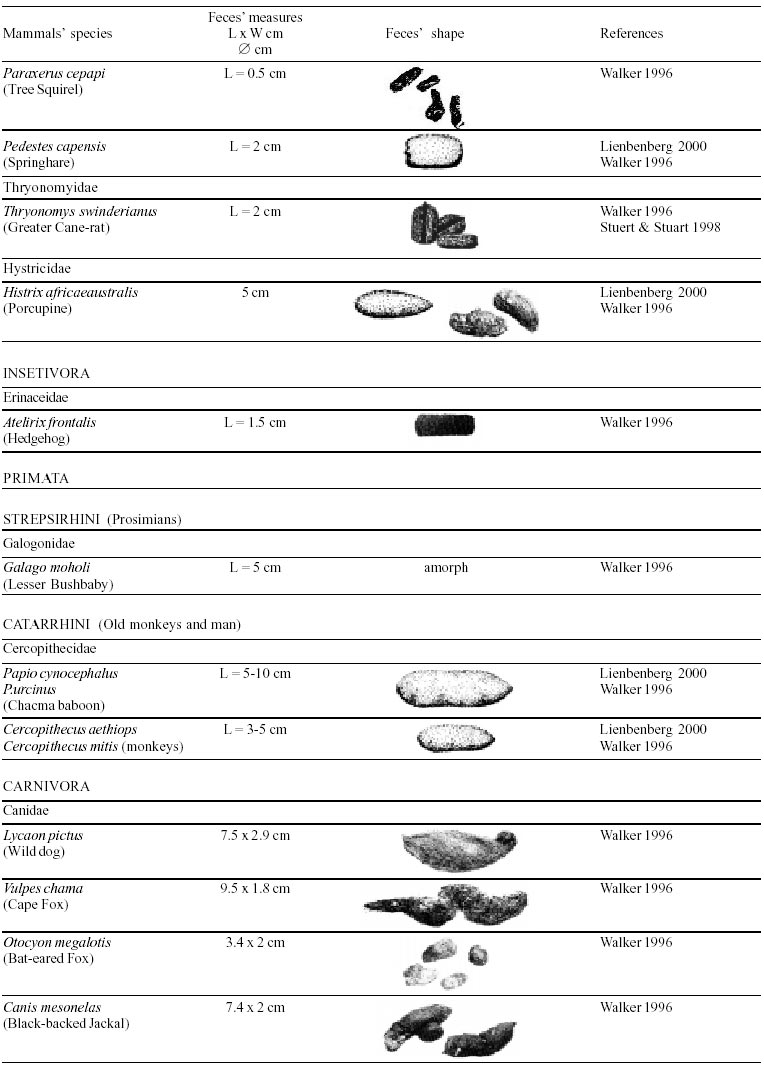
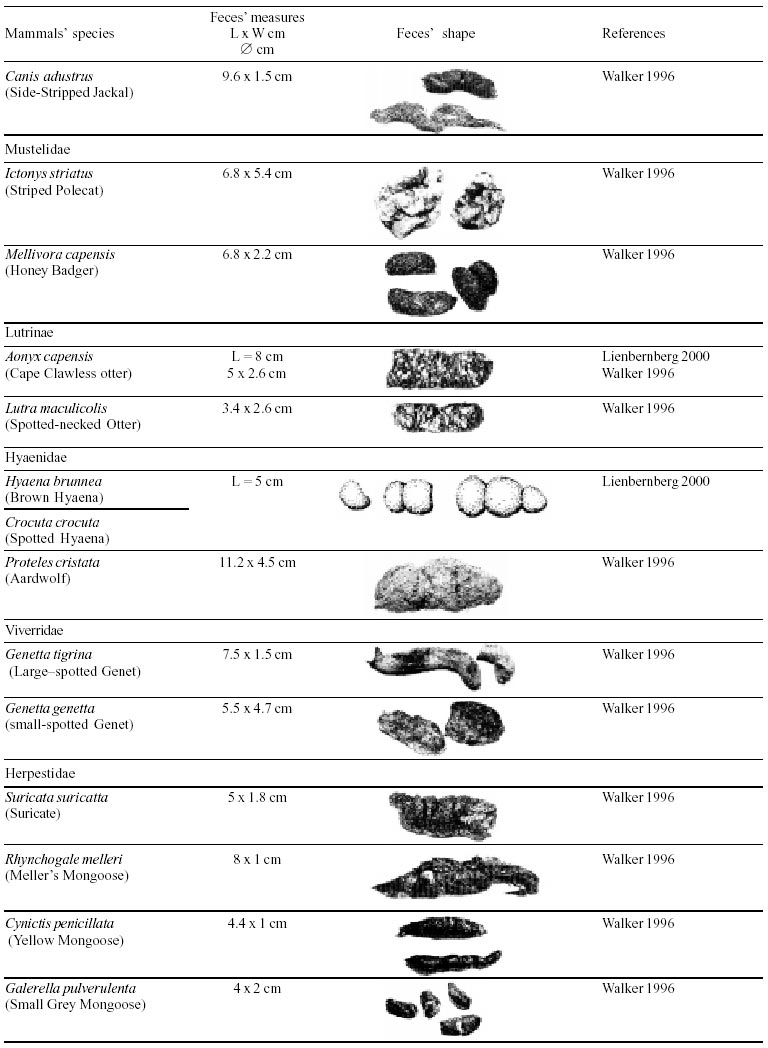
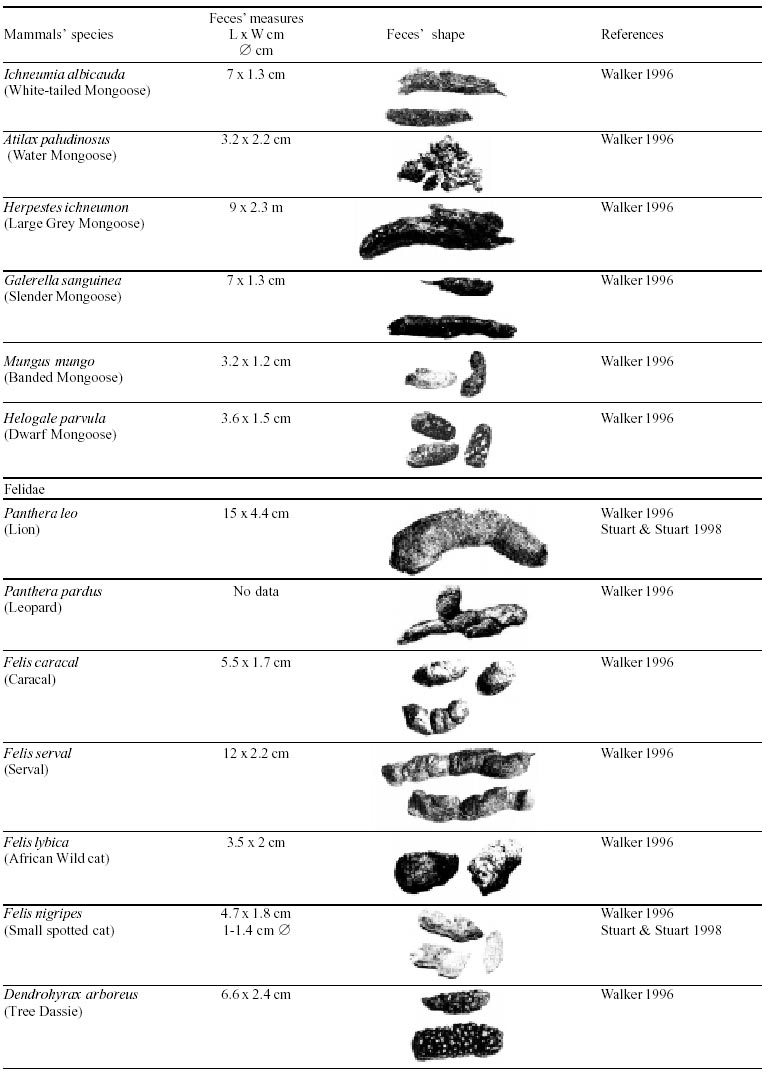
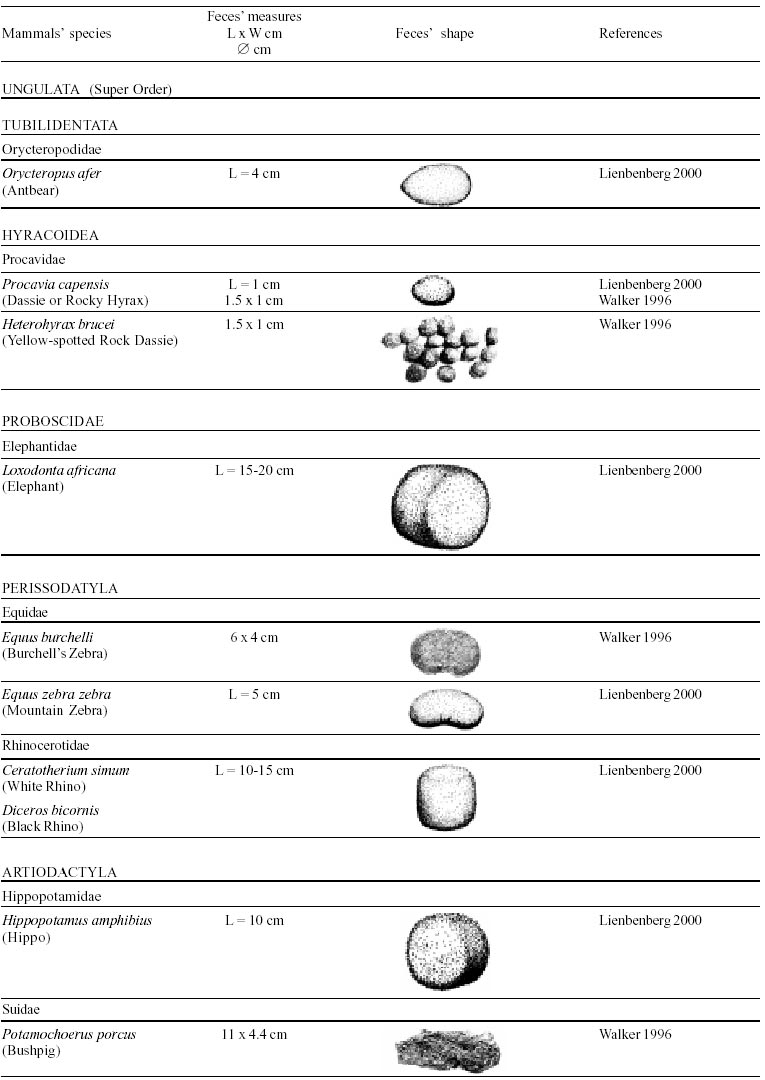
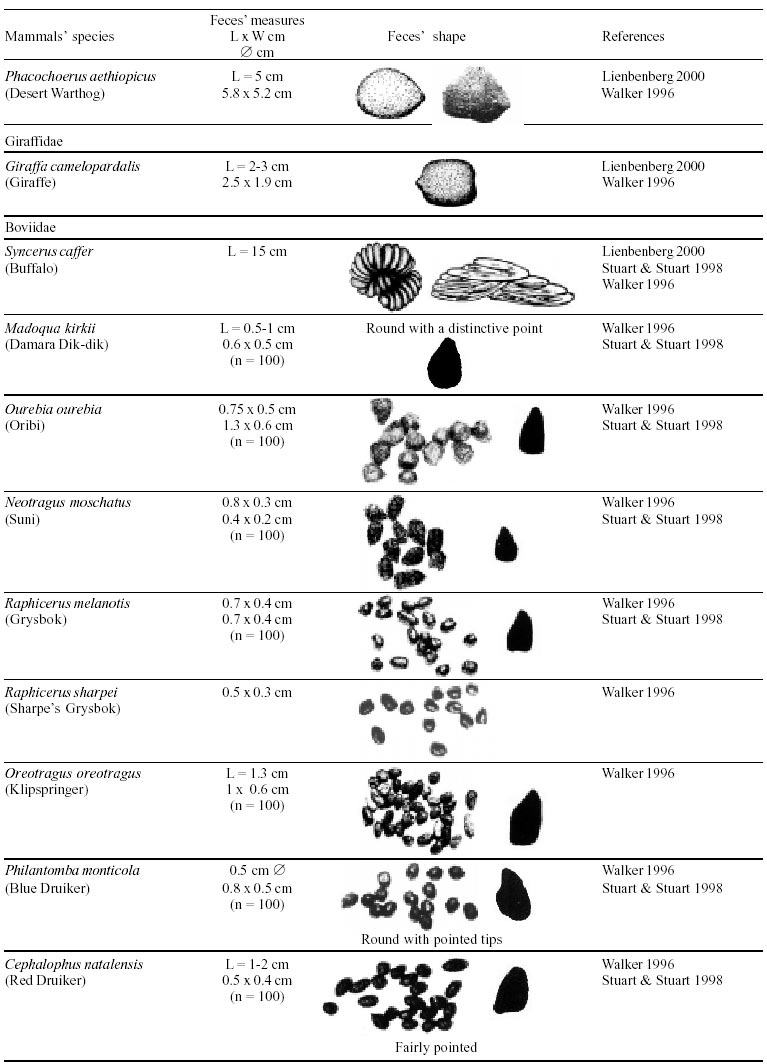
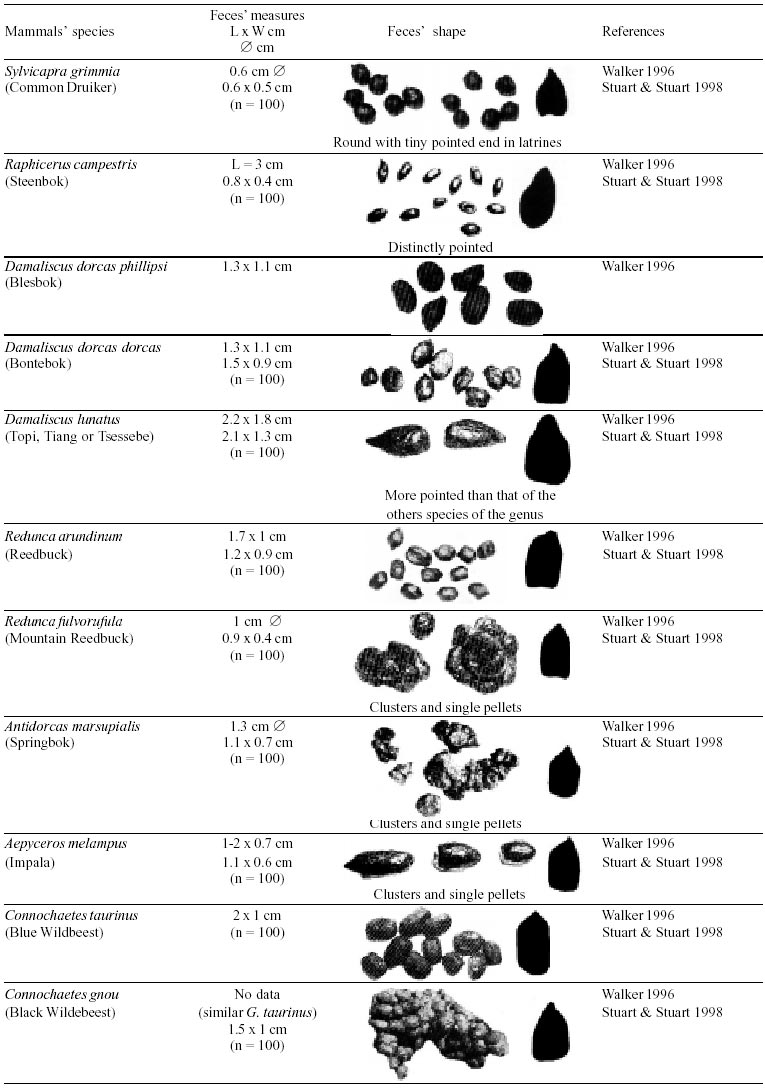
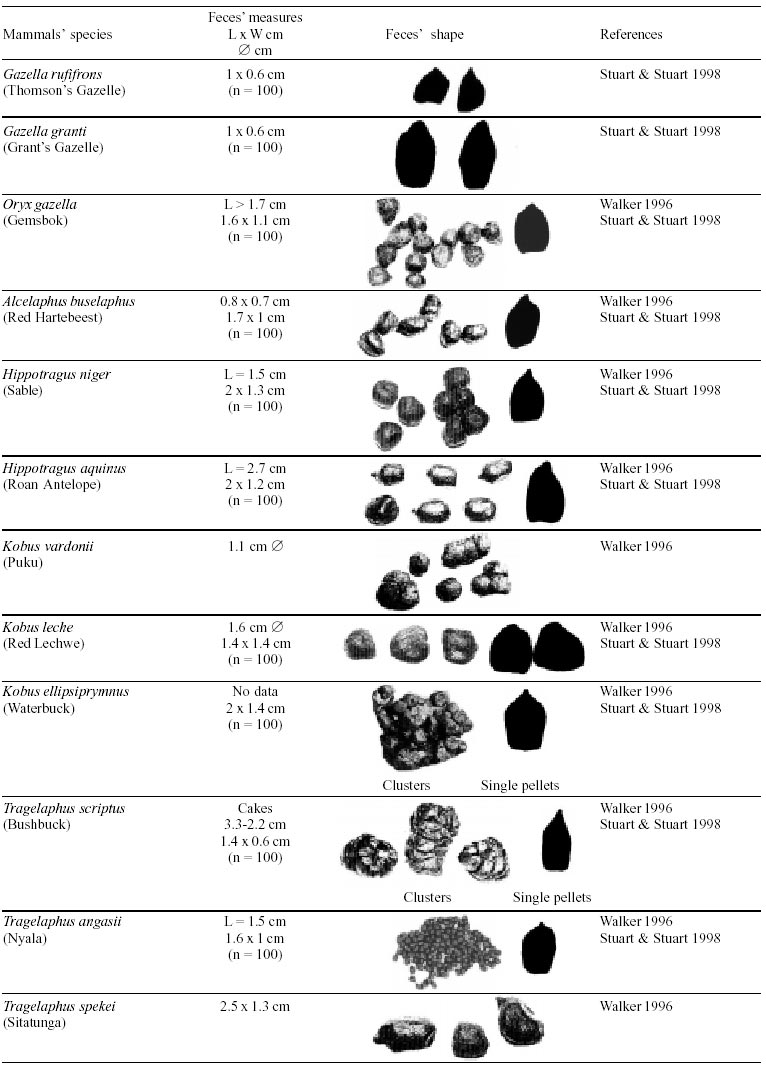
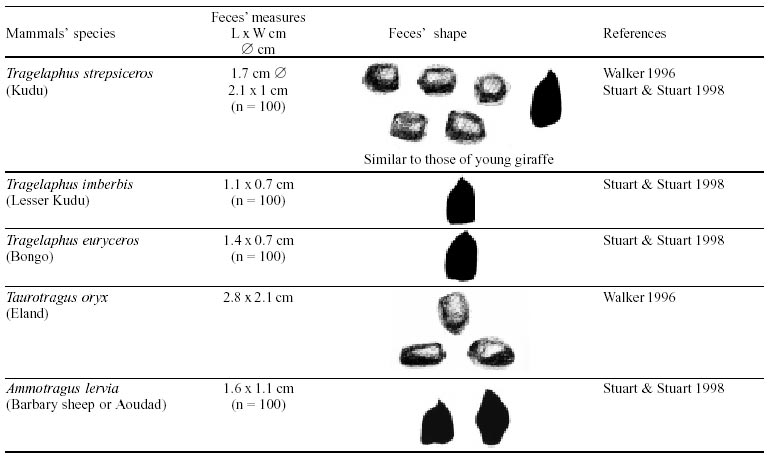
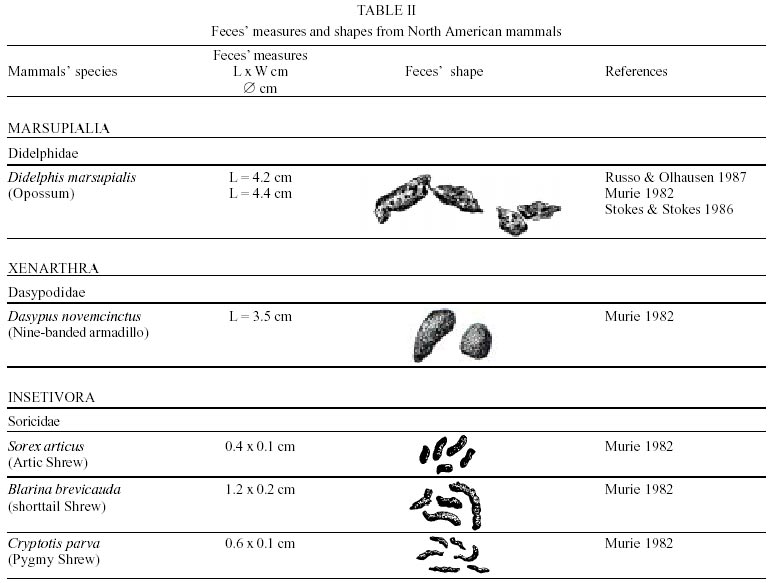
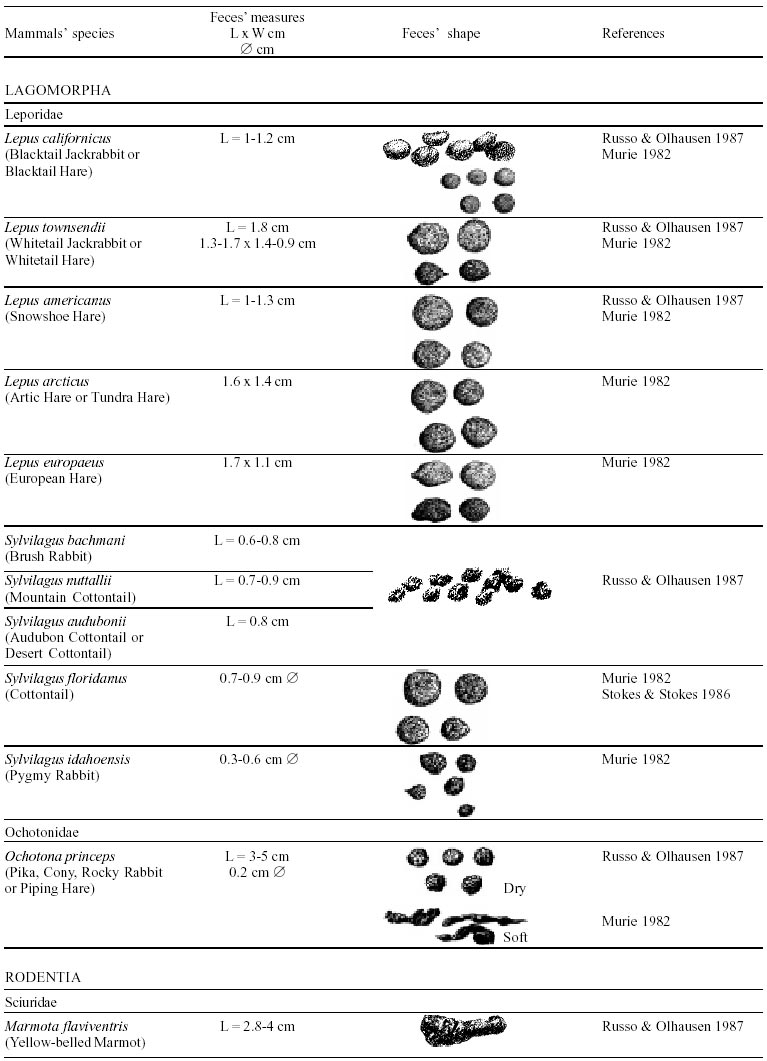
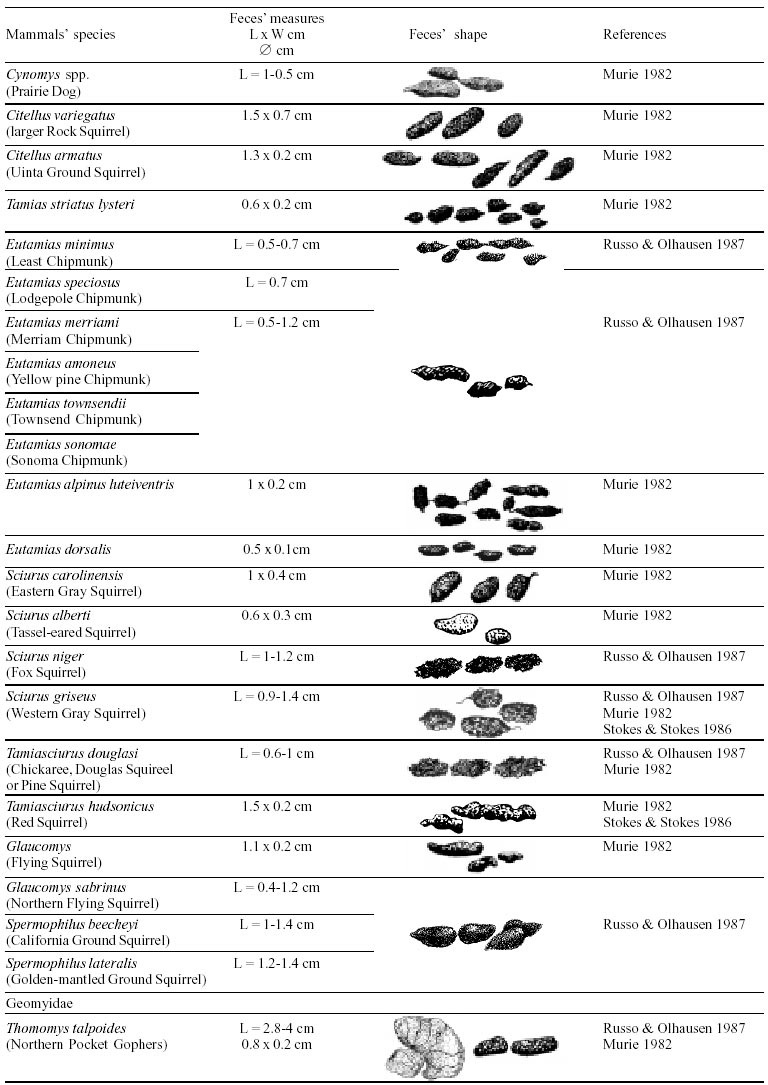
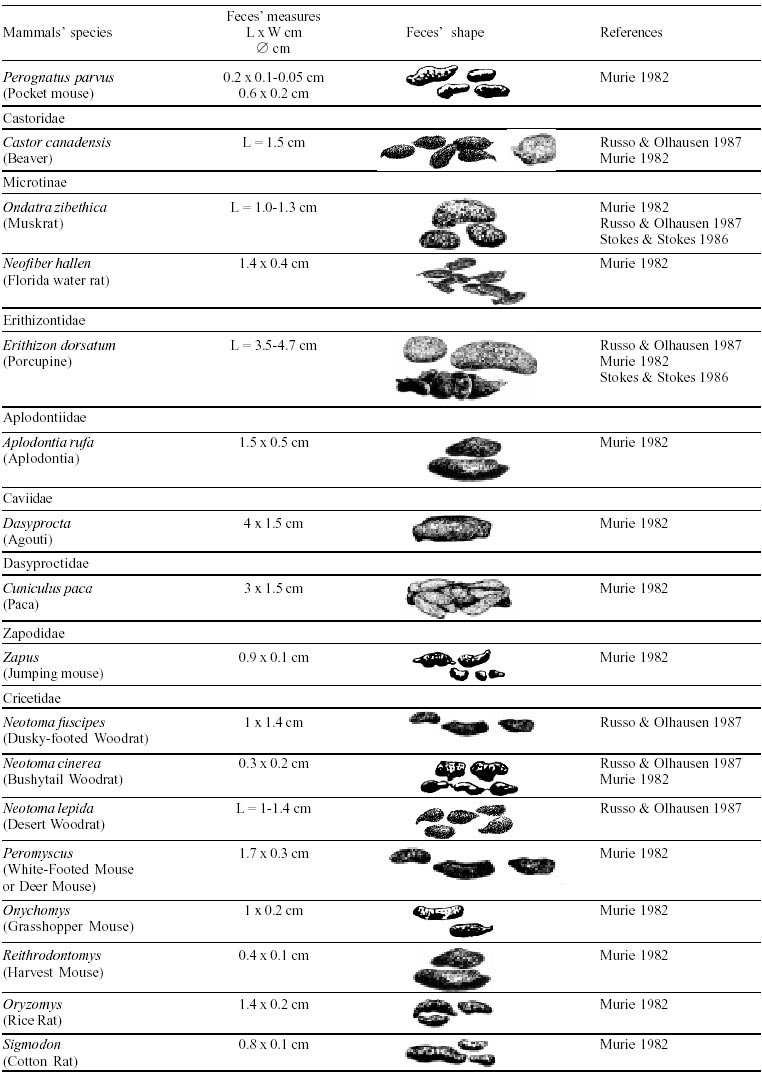
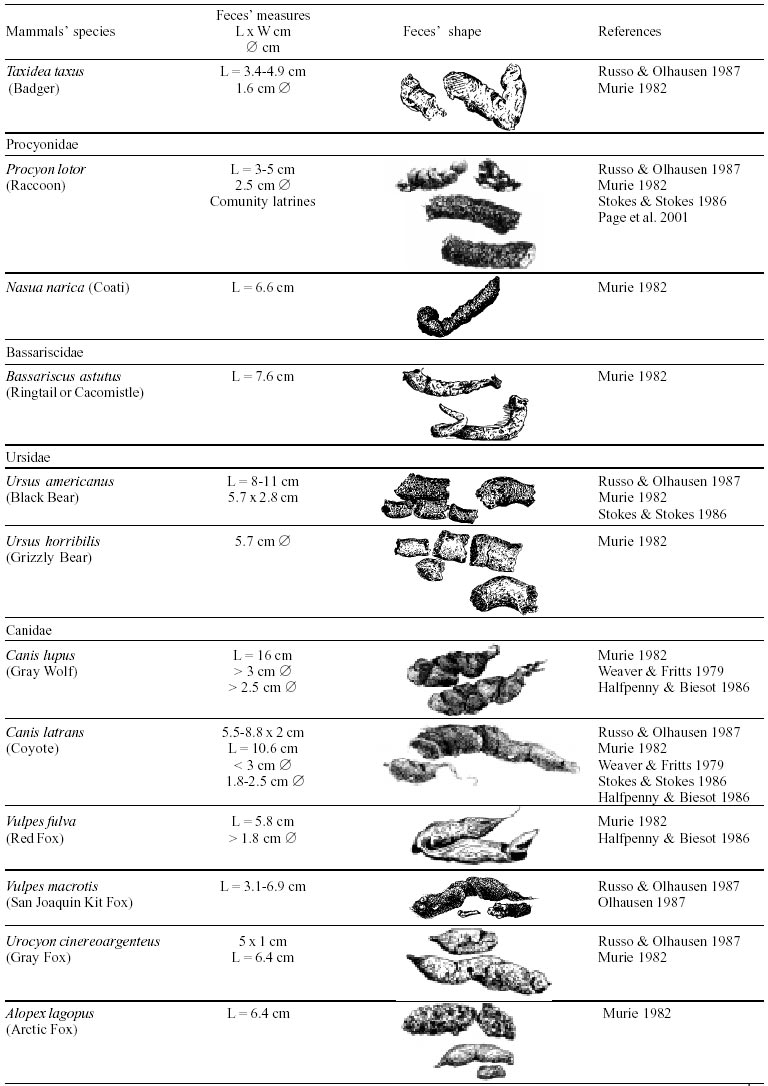
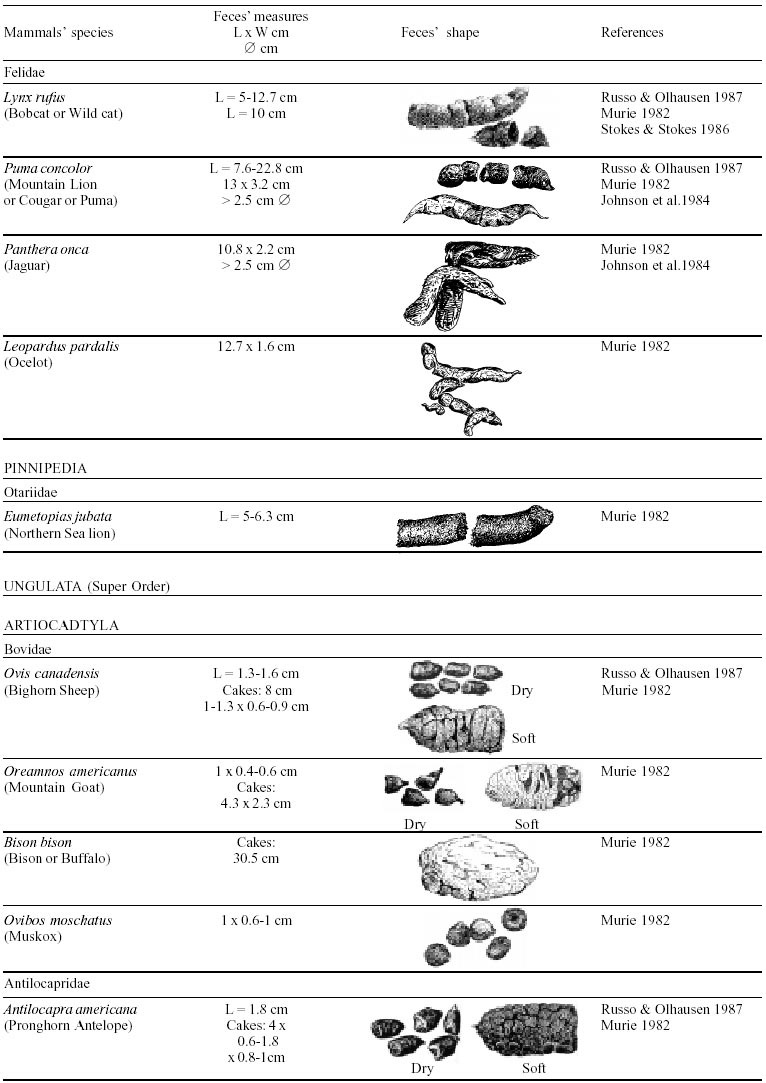
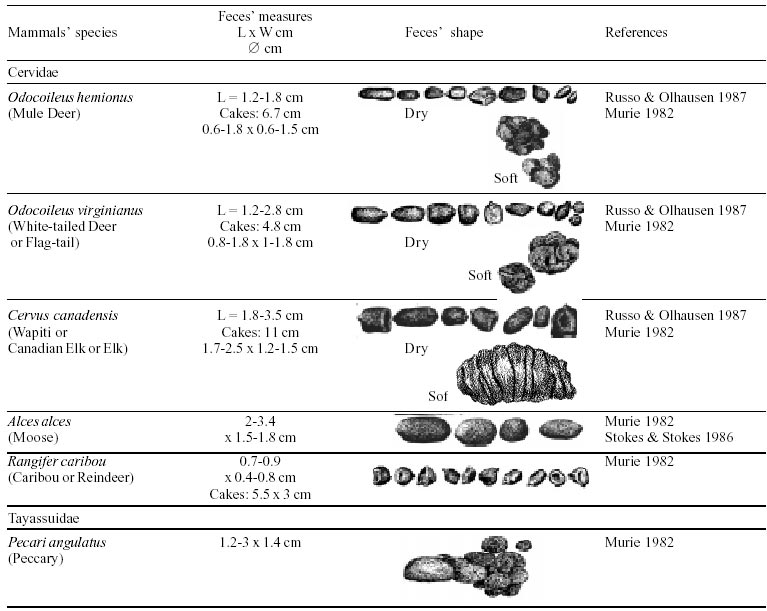
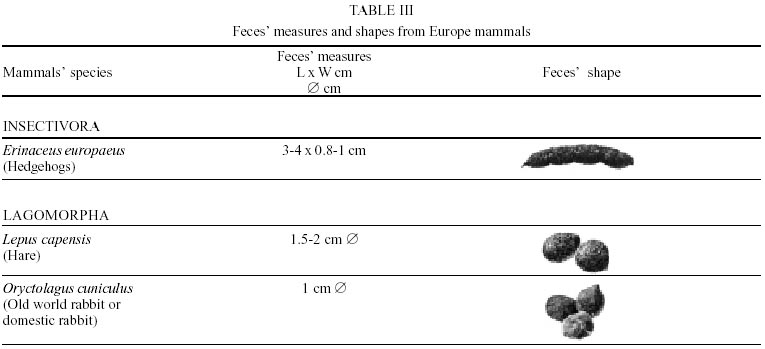
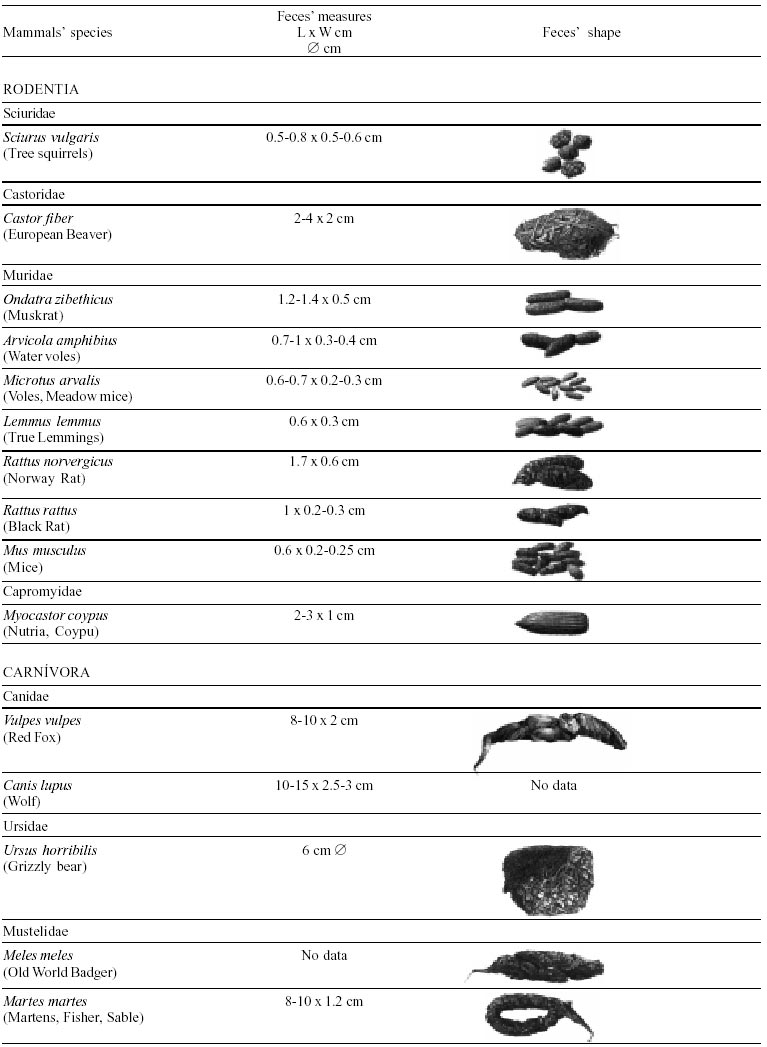
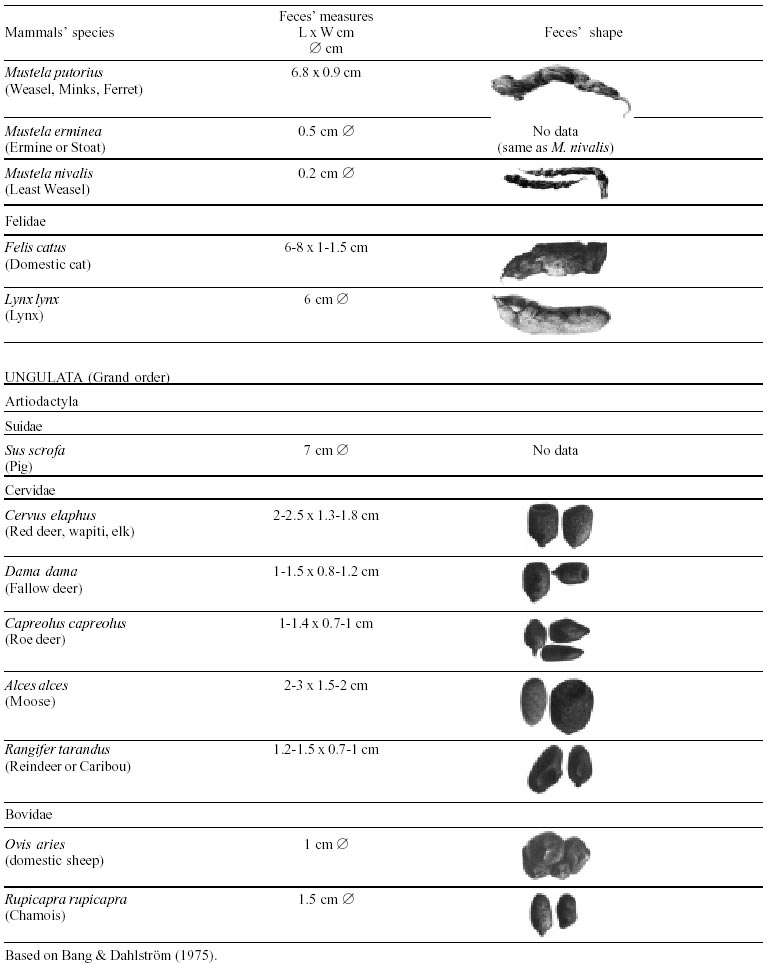
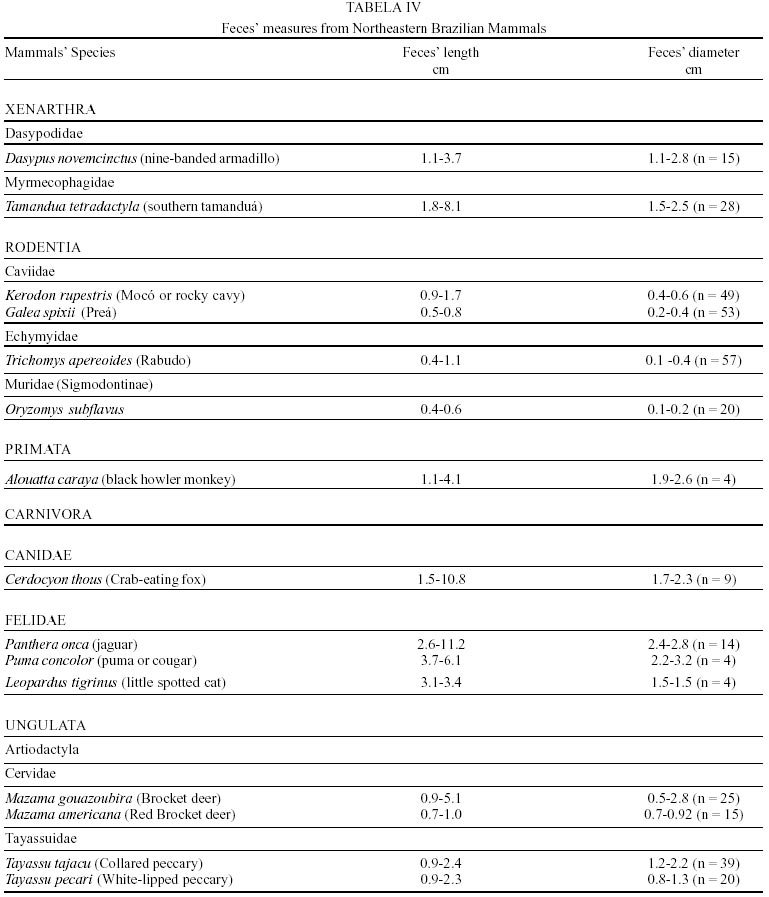
Besides the observer abilities, some other factors influence the find of marks and signs left by animals. Soil characteristics, vegetation, and local climate determine sign and mark conditions. Sandy and loamy soils preserve better footprints than soft soils with thick organic material of superposed layers, and stony areas. However, stony areas, dry ecosystems, and frozen ecosystems provide the best preservation for feces (Bang & Dahlström 1975). On the other hand, it is not easy to identify signs in areas where animals have high demographic densities. Feces are the most evident and most easily recognizable sign (Liebenberg 2000). However, the rarity of some species difficult the observation, as well as others factors like, the presence of buried feces or the behaviour of defecating inside the water or on the branches of the trees. For identification, the original fecal shape must be maintained. Several factors can corrode the original fecal shape through time. These factors include heat, desiccation, or fast decomposition in humid and rainy regions. Fragmentation by other animals such as dung beetles and termites, which frequently consume herbivorous feces, is also another factor that prevents fecal preservation (Stuart & Stuart 1998). Feces can also be consumed by carnivores. For example spotted hyenas eat lion dung and fresh wild dog dung (Stuart & Stuart 1998). Droppings consist of partly digested material and undigested parts of animals and plants. Fecal components may include feathers, bones, teeth, claws, scales, arthropod chitin, seeds and plant tissues, pollen grains, as well as mucus, cells, and a significant amount of living and dead bacteria (Bang & Dahlström 1975, Bjune 2000). Mammal feces have a social communication role (Gorman & Trowbridge 1989). When randomly deposited they show the individual or group home-range, as among American marsupials, lagomorphs, some ungulates, some rodents, and primates. They are used as territorial marks when deposited in small volumes in prominent places such as trail junctions, rocks, trunks, or termite nests. Feces are used as strategic sensorial marks by all Carnivora family species except Hyaenidae (Gorman & Trowbridge 1989, Estes 1991, Romo 1995, Aragona & Setz 2001). Some mammals defecate in discreet individual latrines as do hyaenas (Gorman & Trowbridge 1989) and collared anteaters (Tamandua tetradactyla, Myrmecophagidae : Edentata) in the Brazilian Northeast (Chame 1988). Collective latrines are used by some ungulates, such as antelopes (Walker 1996), Hyrax spp (Kingdon 2001), and some procyonids (Page et al. 2001). Collective latrines can also be used for generations as in the case of Kerodon rupestris (Caviidae: Rodentia), an endemic species of the Brazilian semi-arid region. Feces from this animal are found in rock-shelters and in the archaeological sites of the Serra da Capivara National Park (Ferreira et al. 1991, Araújo et al. 1993). In the carnivores, the secretion produced by the anal gland adheres to the feces during defecation. The secretion of each species has a characteristic and complex odour and it supplies intra and interspecific information of an individual's territory, sex, reproductive state, and movements (Gorman & Trowbridge 1989). The size and the amount of feces produced by each individual varies with age, the type of ingested food, and its absorption capacity (Bang & Dahlström 1975). Size variation is more frequent among herbivores because of the alteration in the quality and amount of food ingested in different seasons. Size varies less among carnivores (Stuart & Stuart, 1998). Food characteristics also affect fecal consistency. Fibrous plants may be the only food found during dry periods or in arid environments, so animals produce hard and more compact feces. During rainy periods or in tropical rainforest ecosystems, the larger consumption of green leaves, sprouts, and fruits produce soft, large, and aggregated feces. Scatology is the science that studies feces (Seton, 1925) and since 1970s the number of studies in this area is increasing (Putman 1984, Halfpenny & Biesiot 1986, Kohn 1997). Several types of information can be obtained from feces and their contents, including the identification of the animal (Seton 1925, Camardella et al. 2000), their activity centers (Walker 1996), diet composition (Johnson & Hansen 1978, Johnson & Aldred 1982, Emmons 1987, 1997, Inagaki & Tsukahara 1993, Chinchilla 1997, Santos & Hartz 1999, Kauhala & Auniola 2001), seasonal diet changes (Corn & Warren 1985, Aragona & Setz 2001), inventory of prey species (Floyd et al. 1978, Emmons, 1987, Camardella et al. 2000), the role of seed dispersion (Fragoso & Huffman 2000, Williams et al. 2000), health condition, and potential entero-parasitosis dynamics (Patton et al. 1986, Page et al. 2001). Researchers have been using feces counting methods for population estimation (Neff 1968). However, the effectiveness of this methodology is still controversial due to seasonal variation, the difficulty in estimating a daily defecation output, and predicting the time of fecal decomposition (Lancia et al. 1996, Patterson 1998). Scatology developed from basic morphometric description to more sophisticated chemical analyses (Nagy & Gilbert 1968, Johnson & Carey 1979, Weaver & Fritts 1979, Weaver & Hoffman 1979, Major et al. 1980, Danner & Dodd 1982, Rollings et al. 1984, Fernández et al. 1997). Recently, the application of molecular biology techniques to the study of feces allowed new approaches for the management of threatened species (Reed et al. 1997). Through DNA recovered and identified from feces, it is possible to distinguish similar feces of sympatric carnivores (Farrel et al. 2000), population variation and biogeography of isolated ape groups in fragmented forests (Jensen-Seaman & Kidd 2001), and the variation in food behavior among individuals of the same species (Fedriani & Kohn 2001). Despite biotechnological progress, the basic initial diagnosis provided by morphometry and the assemblage of signs observed in the field determine the starting point for subsequent studies, and the choice of more sophisticated techniques. For paleoparasitological studies it is important to identify the zoological origin of coprolites. So, the study of fecal morphology is important for modern wildlife study and paleoecological coprolite study. Presented here is a comprehensive summary of fecal morphometric data from bibliographical sources for conspicuous species of terrestrial mammals. These data are from North America, Central America and South America, Europe, and Southern Africa, and Eastern Africa. It also includes the results of my studies in Brazilian arid Northeast. MORPHOMETRIC CHARACTERISTICS OF TERRESTRIAL MAMMAL FECES Data presented here were obtained from scientific journals and also from field guides published by non-academic editors (Burt & Grossenheider 1973, Murie 1974, Bang & Dahlström 1975, Russo & Olhausen 1987, Estes 1991, Walker 1996, Stuart & Stuart 1996, 998, Lienbeberg 2000, Kingdon 2001). The compiled data, presented in Tables I-IV and in the Figure allows the morphometric analysis and the identification of 9 similar fecal groups. Our groups are in accordance with Seton (1925) who emphasized that the form and the contents of feces are excellent guides for the diagnosis of each mammalian Order. They reflect their peculiar anatomy. Feces have a low value to the diagnosis at the Family level, and none at generic level. Seton (1925) presents the curious and seemingly contradictory statement that fecal shape is a valuable and auxiliary consideration in specific diagnosis, and that size and food contents can sometimes separate close related species (Chame 1991). GROUP I
The Felidae family feces can be identified by their compact form with well defined segments and one of the extremities especially tapered (Table I (a, b, c, d, e, f, g, h, i), II (a, b, c, d, e, f, g, h), III (a, b, c) and Figure). Other families and subfamilies can be distinguished by the difference of diet remains, such as in pinnipeds and aquatic mustelids (Lutrinae), whose feces are only composed of fish, crustacean, and mollusc remains. Felid feces reflect strictly carnivorous diet. However, grass leaves ingested to aid hair elimination are also found. Fruit, seed, insect, crustacean, plant tissues, and shell fragments are commonly found in omnivorous canids, mustelids, viverids, and procyonids feces. In North America, felid feces with diameters larger than 2.5 cm can be identified to jaguar (Panthera onca) and puma (Puma concolor). Feces with smaller diameters are assigned to other felid species (Johnson et al. 1984). In Brazilian Northeast a diameter larger than 2.1 cm is enough to separate the great felids (P. onca and P. concolor) from the small felids (Leopardus tigrinus, Leopardus wiedii, Leopardus pardalis, and Herpailurus yaguaroundi) (P < 0.0001, Chame 1988). Morphometric patterns can not distinguish puma from jaguar feces, and both species are sympatric (Emmons 1997). In Costa Rica, Chinchilla (1997) showed that ocelot feces (Leopardus pardalis) have a significantly smaller diameter (c = 2.26 ± 2.46 cm, n = 15) than jaguar (c = 3.15 ± 1.82 cm, n = 16) and puma (c = 2.92 ± 1.09 cm, n = 9) (P < 0.05). As in the other regions of the Americas, the measurements do not distinguish the great feline feces. In similar studies in Peru, Romo (1995) found that the puma fecal diameter exceeded 2.5 cm, whereas Andean Fox (Pseudalopex culpaeus) varied from 1.7 to 2.2 cm, and mountain cat feces (Leopardus colocolo) varied from 1.3 to 1.6cm. Studies of sympatric North American canids show that feces with diameters larger than 3 cm can be identified as of wolf (Canis lupus), and smaller, similarly shaped feces were from coyote (Canis latrans). Only 4.9% of coyote feces are misdiagnosed as wolf feces (Waever & Fritts 1979). However, with chemical and molecular analysis it is possible to identify coyote, fox and bobcat feces (Stokes & Stokes 1986). In Europe, the Gray Wolf feces (Canis lupus) vary from 10-15 x 2.5-3 cm and the Red Fox feces (Vulpes vulpes) from 8-10 x 2 cm (Table III (a, b, c)). Thus, feces from these animals can be identified by morphometry (Bang & Dahlström 1975). In Brazil the maned-wolf (Chrysocyon brachyurus) is the larger canid species. Its feces have diameters larger than 2.5 cm, and also an characteristic odour and texture. Furthermore, its feces contain fruit remains which are distinctive to this animal (Motta-Júnior et al. 1999, Aragona & Setz 2001). In central Brazil (Serra da Canastra National Park) we found a sample of maned-wolf feces with a diameter of 4.5 cm, considerably larger than cougar and jaguar feces. Other Brazilian canids are smaller, so it is easy to identify maned-wolf feces (Dalponte 1997) (Table IV). In Africa the great diversity of medium and large carnivores do not allow a feces morphometric diagnosis criteria (Table I (a, b, c, d, e, f, g, h, i)). Feces of large carnivores can sometimes be totally white as happens with jaguar and puma in America, lion (Panthera leo) and hyenas in Africa, and wolf (Canis lupus) in Europe and North America. White feces are a result of high calcium content as a consequence of bone ingestion (Bang & Dahlström 1975, Chame 1991). They can be also be completely black as lion feces, due to the great amount of blood ingested (Lienbenberg 2000). GROUP II
In this group, shape and diet do not allow to identify the origin of the feces, except for Orycteropus afer (Aardvark) (Table I (a, b, c, d, e, f, g, h, i)), a termite, ant, and larva consumer. GROUP III
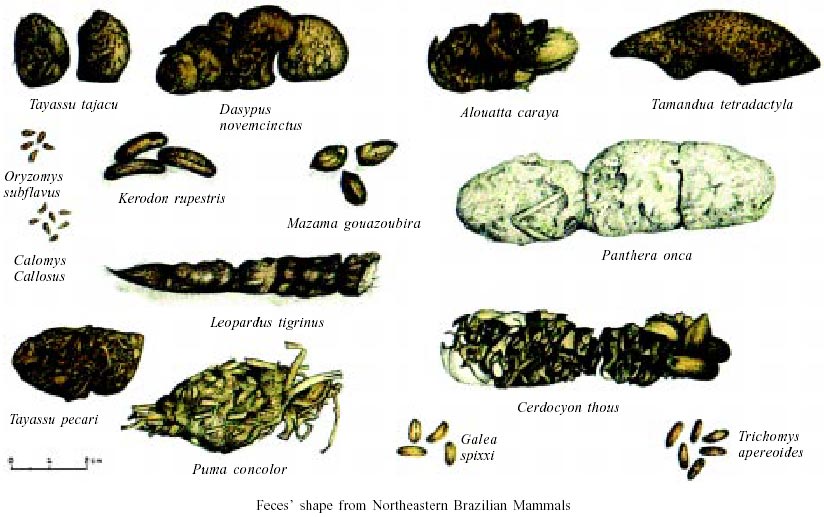
In Northeastern Brazil, feces of Trichomys apereoides (Echymyidae), Oryzomys subflavus (Muridae: Sigmodon-tinae), Calomys callosus (Cricetidae), Galea spixi, and Kerodon rupestris (Caviidae) (P < 0.0001) (Table IV) can be distinguished by diameter measurements (Chame 1988). GROUP IV
The feces of K. rupestris and G. spixii can be diagnosed accurately. K. rupestris feces present prominent furrow in the concave face and a diameter larger than 0.8 cm. In G. spixii feces the furrow is in the convex part and the diameter is smaller than 0.8 cm (Table IV, Figure). GROUP V
The feces of two deer species from Brazilian semi-arid region (Mazama gouazoubira and Mazama americana) cannot be differentiated by the shape and size (Table I (a, b, c, d, e, f, g, h, i), II (a, b, c, d, e, f, g, h), III (a, b, c)), as observed also in similar species in Africa, North America and Europe. GROUP VI
GROUP VII
GROUP VIII
This group also includes South American anteaters (Myrmecophagidae: Xenarthra). Collared anteater feces (Tamandua tetradactyla) are cylindrical and continuous. They measure 1.8-8.1 x 1.1-2.8 cm and easily break when they fall on the ground. They are deposited close to shelters, in easily recognisable individual latrines. The surface is flat and ant and termite remains can be observed with naked eye. Although there are no morphometric studies of feces of other species of this family, it is possible that their size could be used to distinguish them. Giant anteater (Myrmecophaga tridactyla) is the larger species weighing from 22 to 39 kg. The silky or pygmy anteater (Cyclopes didactylus) just weighs between 155 to 275 g. Two other species have the same size but do not share the same geographical distribution (Emmons 1997). These species are T. tetradactyla (that weighs from 3.6 to 8.4 kg) and the Northern tamandua (Tamandua mexicana). GROUP IX
Usually, primate feces can be identified if the feeding sites are known such as with capuchin monkey studies (Cebus apella, Cebidae) in southern Brazil (Pizo & Oliveira 1999), and black-howler-monkeys (Alouatta caraya, Atelidae) in the Brazilian Northeast (Chame & Olmos 1997). FINAL CONSIDERATIONS
Nine groups can be characterised by fecal morphometry, and the patterns of shape and size of the terrestrial mammal feces are sufficiently consistent to group them (Bang & Dahlström 1975). Although the size of the feces varies with individual animal age, as well as food habits, a size limit can be standardised and attributed to them. Particularly, in my studies conducted in the Northeast of Brazil, the shape and diameter of the feces are a better specific indicators than length. Shape can be identified as the first indicator for the diagnosis of the fecal origin, cor roborating the statements of Seton (1925). The statistical analysis of measurements can distinguish species of the same group, as in rodents of the Northeast of Brazil (Chame 1988). When the diet is well known for a determined area, it can be an important factor to distinguish species, as in the case of the African rhinoceros and South American canids (Chame 1991, Motta-Júnior et al. 1996, Dalponte 1997, Aragona & Setz 2001). The groups of feces identified in this work suggest that the morphology of the feces may reflect a species phylogeny, corroborating paleontological data about the evolution and radiation of the mammals. It is expected that the definition of morphometric patterns to identify terrestrial mammal feces can be used not only for the progress of field studies of current fauna, but also to stimulate biomolecular studies based on feces for paleoecological, and paleoparasitological studies (Chame et al. 1991, Araújo et al. 2000) that use coprolites as a primary source of investigation. As the study of the feces and coprolites starts in the field, with the tracking animals or with the archaeological excavations, it is the observer's acuity and sharpness in the gathering the largest amount of information left by the animals in nature, together with laboratory results, that makes it possible to rebuild movements, ecosystems, and biological and ecological relationships. In essence, it allow us to determine what has happened when there were no observers present. ACKNOWLEDGMENTS
To Gabriela Landau Remy for providing the African Field Guides. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03014t2f.jpg] [oc03014t2h.jpg] [oc03014t1d.jpg] [oc03014t4.jpg] [oc03014t2d.jpg] [oc03014f1.jpg] [oc03014t2b.jpg] [oc03014t1i.jpg] [oc03014t1b.jpg] [oc03014t2g.jpg] [oc03014t2a.jpg] [oc03014t1f.jpg] [oc03014t2e.jpg] [oc03014t3c.jpg] [oc03014t2c.jpg] [oc03014t1c.jpg] [oc03014t1e.jpg] [oc03014t3a.jpg] [oc03014t3b.jpg] [oc03014t1g.jpg] [oc03014t1a.jpg] [oc03014t1h.jpg] |
| |||||||||

 Cylindrical
or rounded pellets usually pointed at one end and concave in the other extremity
(Table I (
Cylindrical
or rounded pellets usually pointed at one end and concave in the other extremity
(Table I ( Big
and cylindrical feces like bars that characterise large ungulates such as the
elephants, hippopotamus, and rhinoceros. The two African species of rhinoceros
use collective latrines which can be shared when they are in the same territory
(Stuart & Stuart 1998). Dung of the white rhinoceros (Ceratotherium simum)
contains only grass, while black rhinoceros (Dicerus bicornis) is easily
identified by its fibrous and woody contents (Walker 1996, Liebenberg 2000).
Big
and cylindrical feces like bars that characterise large ungulates such as the
elephants, hippopotamus, and rhinoceros. The two African species of rhinoceros
use collective latrines which can be shared when they are in the same territory
(Stuart & Stuart 1998). Dung of the white rhinoceros (Ceratotherium simum)
contains only grass, while black rhinoceros (Dicerus bicornis) is easily
identified by its fibrous and woody contents (Walker 1996, Liebenberg 2000).
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}