 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98(Suppl. I), January 2003, pp. 195-205 Pathoecology of Chiribaya Parasitism Elizabeth Martinson/+, Karl J Reinhard*, Jane E Buikstra**, Katharina Dittmar de la Cruz***
P. O. Box 85028, Fairbanks, AK, USA
*School of Natural Resource Sciences, University of Nebraska-Lincoln, USA **Department
of Anthropology University of New Mexico, Albuquerque, USA ***Department of
Zoology, Insect Genetics Lab Brigham Young University, Provo, UT, USA Received 26 August 2002 Code Number: oc03029
The excavations of Chiribaya culture sites in the Osmore drainage of southern Peru focused on the recovery of information about prehistoric disease, including parasitism. The archaeologists excavated human, dog, guinea pig, and llama mummies. These mummies were analyzed for internal and external parasites. The results of the analysis and reconstruction of prehistoric life from the excavations allows us to interpret the pathoecology of the Chiribaya culture.
Key words: pathoecology - archaeoparasitology - endoparasitism - ectoparasitism - Peru Pathoecology is the study of the biotic, abiotic, and cultural environments of disease. In the archaeological setting, the pathoecological interpretation of parasite data is based on reconstruction of these environments. Presented here is the pathoecological reconstruction of the Chiribaya culture of Southern Peru. The excavations of Chiribaya sites, done in the 1990s, were conducted, in part, to recover evidence of parasitic disease. This was the first time several sites from the same archaeological culture were excavated for the comparative study of ancient parasitism. The funding for the excavations and analysis came from National Science Foundation grant Grant BNS89-207691990-1993 "Chiribaya: A Biocultural Approach to the Study of a Prehistoric Andean Polity" awarded to Buikstra. This was an important historical marker for the then fledgling field of ar-chaeoparasitology because interpretation of the excavated sites allowed us to place parasitic disease into a cultural framework. The archaeoparasitology of the Chiribaya is complex and was eventually split into four separate research lines. The louse ectoparasitism of humans was unusually rich and deserved independent treatment (Reinhard & Buikstra, this volume). The parasitism of domestic guinea pigs became a special interest for Katharina Dittmar de la Cruz (Dittmar et al. 2000, this volume, Dittmar de la Cruz 2001). The endoparasitism and ectoparasitism of domestic camelids became a second line of enquiry (Leguía & Casas 1999). Finally, human endoparasitism was a fourth line of enquiry and is the primary subject of this paper. It is our purpose to present a general overview of Chiribaya archaeology and then summarize the parasite ecology of the drainage. THE PHYSICAL SETTING
The sites studied are located within the Osmore drainage of far southern coastal Peru (Fig. 1). The Osmore drainage is located within the Department of Moquegua between 70"27' and 71"20' west longitude, and 16"52' and 17"42' south latitude (Rice 1989). The drainage rises from sea level to 5,100 m above sea level in just 139 km (Burgess 1999). This extreme elevation change leads to marked disparities in climate between areas that, on the horizontal plane, are not far removed from one another. The Osmore drainage is a deeply incised, steep-walled valley. The flood plain is generally very narrow, and the hills surrounding the floodplain are often steep and cut with quebradas (gullies). Irrigation is compulsory in the lower and mid valleys since there is negligible rainfall (Rice 1989). Agricultural goods were grown in the river valley prehistorically, and there is evidence of camelid herding. The Chiribaya also kept cuyes (guinea pigs). Wild plants and animals could be found along the river and in the lomas (fog gardens) that occur on the coast. The Pacific Ocean would have provided the Chiribaya with a variety of fish, marine mammals, sea birds, crustaceans, and mollusks. The outstanding characteristics of the lower and middle part of the Osmore drainage are extreme aridity and a virtual absence of vegetation. The town of Ilo, located on the coast, receives an average of only 5.3 mm of precipitation annually, with most of it in the form of fog (Rice 1989). Breaking the monotony of the extreme aridity are El Niño events. El Niños occur on average of once every 7 years (Moseley 1975). During these events, the coast is inundated with rain, and the fishery is disturbed by the changes that occur in the Humboldt Current, which parallels the coast. El Niños can last from 6 to 18 months (Thayer & Barber 1984) and the effects can be devastating. In prehistoric communities reliant on resources from the sea, the effects of an El Niño could have been tremendous, greatly reducing or eliminating a predictable and plentiful calorie and protein source. The torrential rain that accompanies El Niño events can cause major flooding that destroys irrigation canals, farmland, and settlements, including any stored food. El Niños tend to strike a considerable length of the coastline, so coastal neighbors can not be relied upon for help. In addition, the high altitude zones undergo a period of drought while the coast is inundated, so aid from highland neighbors is not an option either (Sutter 1997). Disease can be rampant in the aftermath of an El Niño, due to the presence of stagnant water, the lack of clean water sources, and damaged maritime and agricultural economic systems (Satterlee 1993). The effects of prehistoric El Niños would no doubt have been even more severe than they are today, due the lack of modern medicine, labor, and technology (Jackson & Stocker 1982, Satterlee 1993). Cabello de Balboa [1951(1586)] notes that ten years after the El Niño of 1576, the Trujillo area had still not recovered completely from the damage done. The climatic regime of the area has created a nearly ideal environment for archaeological preservation. Outside of El Niño events, temperature and humidity are stable, creating an almost ideal environment for the preservation of archaeological materials. Organic preservation in the area is astounding. Botanical remains, textiles, corporal soft and hard tissues, and hair are all very well preserved, and provide an unusually complete record of prehistoric life. The extreme aridity of the area, in general, decreases the likelihood that geohelminths and other fecal-borne helminths such as hookworms and Strongyloides were a problem for the Chiribaya. These parasites require moisture and shade to survive, and these "resources" are in short supply in the Osmore drainage. However, individuals involved with irrigation agriculture would have been at a greater risk for parasitic infection by these species, since helminth eggs and larvae are more likely to have occurred in irrigated fields that were shaded by crops or by the trees that grow in the flood plain. The fact that the river is the only source of water for those individuals living within the drainage suggests that water contaminated with human and animal waste could have played a role in the spread of parasites. It seems reasonable to assume that sites located downstream of large population aggregations were at a greater risk than at those located upstream. THE CULTURAL SETTING
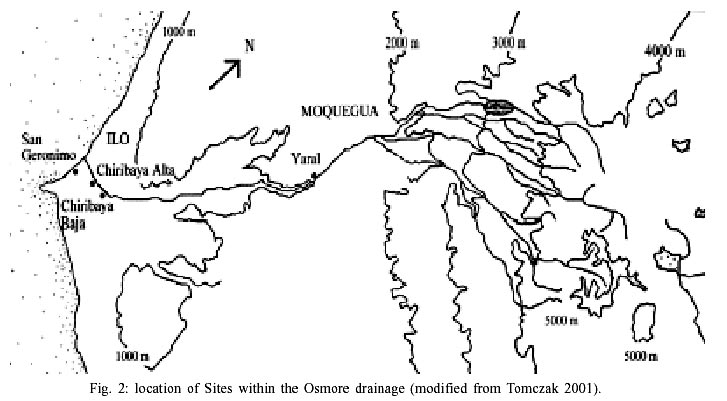
Humberto Ghersi Barrera first defined the Chiribaya culture in 1956. Ghersi excavated seven tombs in the lower Osmore drainage, from which he described distinctive polychrome ceramics with complex abstract geometric designs, and textiles of varied colors with geometric and zoomorphic designs. After Ghersi's (1956) definition of Chiribaya, work on the Chiribaya lingered until 1981, when Belan reported Chiribaya ceramics and textiles from the Ilo, Peru area. Belan (1981) defines the Chiribaya spatial extent as reaching from the Tambo to the Azapa valleys, and temporally he places Chiribaya between AD 1000 and AD 1350. This temporal placement of Chiribaya has been sustained in the literature, with most recent works citing Chiribaya as being a post-Tiwanaku phenomenon, lasting from AD 950/1000-1350 (e.g. Burgess 1999, Clark 1993, Jessup 1990a, Owen 1993, Rice 1993, Stanish & Rice 1989, Sutter 1997). However, a new analysis of radiocarbon dates suggests otherwise (Lozada 1998). The new dates suggest that the Chiribaya culture may have been present in the coastal valley as early as AD 700. Population of the lower valley boomed during the Chiribaya presence (Owen 1993). The population expansion could have promoted some infectious parasitic disease similar to the increase in parasitism found by Reinhard in the deserts of Arizona, Utah and New Mexico when population increased (Reinhard 1988, 1992). Reycraft (1998) argues that the Chiribaya reached their maximum population at approximately AD 1350. This florescence was dramatically cut short by a "mega" El Niño event (Satterlee 1993) that has been dated to approximately AD 1350. This El Niño event is believed to have decimated the agricultural systems of the Chiribaya (Satterlee 1993) and it has been estimated that it reduced human population by as much as 80% (Owen 1993). Chiribaya settlements reach from sea level on the coast to 3000 m above sea level (Jessup 1990a, Stanish 1989), with the majority of sites found between 0 and 1000 m above sea level (Belan 1981). To date, 28 Chiribaya sites and 21 Chiribaya cemeteries have been recorded in the Ilo area (Owen 1993). Six of these sites are located along the coastline, with the remainder of the sites found in the river valley. Sites within the valley focus on both lomas resources and agricultural products, and the coastal sites are more oriented toward marine resources (Buikstra 1995, Burgess 1999, Jessup 1990a,b, Lozada 1998, Sandness 1992). Rectangular compounds located on terraces supported by stone retaining walls characterize Chiribaya architecture (Owen 1998). Typically there is a heavy cane perimeter wall that encloses a series of rooms, storage areas, patios, and corridors (Owen 1998, Rice 1993). The substantial nature of the compounds, evidence of remodeling (Rice 1993), and the amount of midden accumulation at the sites suggests permanency (Owen 1998). The data discussed here are drawn from four Chiribaya sites: Chiribaya Alta, Chiribaya Baja, San Gerónimo, and El Yaral (Fig. 2). A brief description of each of these sites is provided.
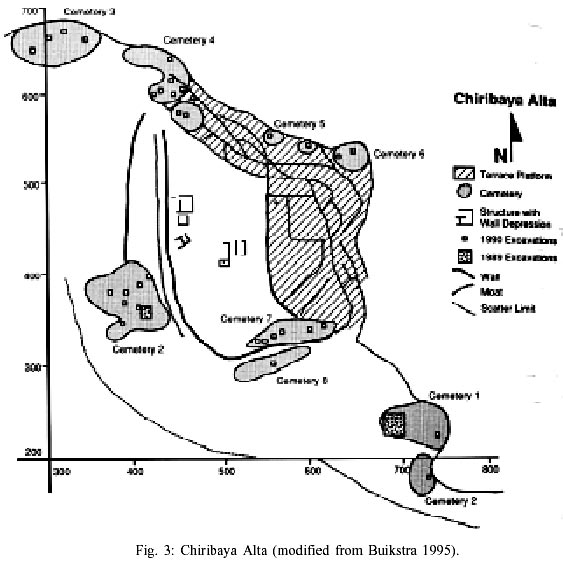
Chiribaya Alta Chiribaya Alta is a 36-hectare site (Buikstra 1995, Rice 1993, cf. Burgess 1999) located approximately 5 km inland of the river mouth (Tomczak 2001). The site has nine cemeteries, a wall and ditch system, residential and agricultural terraces, and large cane-walled structures (Buikstra 1995, Rice 1993) (Fig. 3). Excavations at Chiribaya Alta were undertaken by Jane Buikstra in the late 1980s and early 1990s and focused on mortuary contexts. Chiribaya Alta is unanimously acknowledged as the center of the Chiribaya cultural complex (Owen 1993, Sutter 1997, Burgess 1999). Although the exact nature of Chiribaya Alta's importance is not understood, the characteristics of the site that suggest its centrality are obvious. These characteristics are discussed below. First, the location of Chiribaya Alta is ideal for a central site. Unlike other Chiribaya sites, Chiribaya Alta is not located on the valley bottom. Instead, it is located on the southern edge of the valley, 100 m above the riverbed on the Pampa Inalámbrica. From the north edge of the Pampa Inalámbrica it is a steep drop to the valley floor below (Burgess 1999). This location provides Chiribaya Alta with a commanding view of the valley (Jessup 1990a). Additionally, the site is located near one of the best entries into the valley (Jessup 1990a). Chiribaya Alta's location would have allowed the inhabitants of the site to monitor movement in and out of the valley. Being on the pampa, Chiribaya Alta was also closer to lomas vegetation than were the other sites. The lomas could have been used as a source of food for the inhabitants of Chiribaya Alta, and may have provided forage for camelids. Agricultural land was available immediately below the site along the floodplain, and a variety of marine resources was only 2 km away (Tomczak 2001). It appears that this site was located to both monitor and take advantage of all available resources. Second, the site has evidence of fortification that no other Chiribaya site has. As mentioned above, Chiribaya Alta has a wall and ditch system that separates the site from the surrounding pampa (Belan 1981, Buikstra 1995). Owen (1993: 186) believes that the wall and ditch system was not built for protective purposes. He cites a lack of evidence for warfare among the Chiribaya in his argument (see Burgess 1999). Additionally, there is no internal water source, so the site could not have withstood long sieges. Finally, the site would have only protected a small percentage of the residents of the drainage, and would have been of little use to individuals who lived more than a few kilometers away and could not have taken refuge at the site when necessary. Based on this information, Owen has argued that instead of protecting the site and its inhabitants, the wall and ditch system may have served to separate the site and its inhabitants visually and symbolically from other sites and people in the valley. Chiribaya Alta, with its overbearing location and seclusion from its surroundings, would have created a powerful visual message. No other Chiribaya site seems to have been constructed to send such messages. In contrast to the mortuary contexts, very little is known of the domestic contexts at Chiribaya Alta. Domestic terraces have been mapped, but no systematic excavations have taken place, nor are they likely to occur in the future because of the on-going destruction of the site by looters. Based on the surface evidence, Chiribaya Alta domestic areas are similar to the domestic architecture of Chiribaya Baja and El Yaral (Jessup 1990a), with outlines of multi-roomed rectangular structures (Miranda 1993, Owen 1993, Buikstra 1995) and plaza areas within the confines of the wall and ditch (Rice 1993). In addition, residential terraces line the slope that descends to the valley bottom from the north edge of the site.
San Gerónimo San Gerónimo is located approximately 200 m inland of the mouth of the Osmore River (Jessup 1990b). Rice (1993) places the dimensions of the site at approximately 12 hectares (Fig. 4). The site has previously been referred to as "Boca del Rio" (Belan 1981, Ghersi Barrera 1956), although more recent research has consistently used the name "San Gerónimo". The two available radiocarbon dates from the site are AD 1020 and AD 1156. Jessup (1990b) conducted a rescue project that included the excavation of approximately 90 Chiribaya burials. These burials were found in midden areas associated with other archaeological features, rather than in the formally bounded cemetery areas seen at other Chiribaya sites (Buikstra 1995). The tombs typically contain a single individual placed in a flexed, seated position and wrapped in textiles. Common grave goods include textiles, ceramic vessels, wooden keros, miniature boats, tools for use in textile manufacture, and fishing implements. There are several burials with numerous, finely made objects, suggesting status differentiation similar to that observed at Chiribaya Alta. Little work has been done on the domestic contexts at San Gerónimo. Jessup's (1990b) work at San Gerónimo revealed 18 m2 of prepared floor and numerous storage features, hearths, garbage pits, and significant midden accumulation. In addition, Garcia Márquez (1988) reports the discovery of cane structures at the site. The location of San Gerónimo suggests that marine resources were primary in the site economy, and the archaeological evidence supports this. Many of the males at the site are buried with fishing-related items, including hooks, nets, floaters, weights, and miniature boats. The middens at the site contained great quantities of fish and shellfish (Jessup 1990b). Jessup (1990b) has pointed out that in addition to the resources from the sea, San Gerónimo had an equally rich and diverse terrestrial resource base. The site had access to estuarine and lomas resources, and the large number of camelid remains at the site suggests that they practiced pastoralism. In sum, San Gerónimo was a village with access to numerous rich resources, but the focus was clearly on marine resources. Funerary contexts indicate a mortuary pattern and range of social status similar to that seen at Chiribaya Alta. The location of this site near the river might have exposed its inhabitants to insect- and parasite-born diseases that would not have been a threat to the inhabitants of Chiribaya Alta.
Chiribaya Baja Chiribaya Baja, formerly referred to as "El Algarrobal," is the type-site for the Chiribaya culture (see Ghersi Barrera 1956) (Fig. 5). More recent excavations have been undertaken in the domestic contexts of the site by David Jessup, and in the mortuary contexts by Lozada and Torres (under the direction of Jane Buikstra) and Gerardo Carpio (under the direction of Sonia Guillén). Chiribaya Baja is located approximately 8 km from the coast (Tomcazak 2001, Rice 1993), on the south bank of the river. It is below and within visible range of Chiribaya Alta (Rice 1993). The 14-hectare site consists of residential terraces that extend up the valley wall from the floodplain (Jessup 1990a, Rice 1993). The site is located near the largest tract of irrigable land in the lower valley, and has obvious agricultural ties (Jessup 1990a). Chiribaya Baja was the largest agricultural site in the valley during the Chiribaya presence (Jessup 1990a) and was the apparent focal point of the Chiribaya population (Owen 1993). The mean calibrated date for the site is AD 1025. Miranda (1993) has reported on the domestic contexts at the site. The residential structures consist of rectangular cane and wood structures, placed upon terraces (Rice 1993) supported by foundations of stones. This pattern is similar to that found at the other Chiribaya sites. While the importance of agriculture to the site is evident in the site's location and midden content, Miranda (1993) has pointed out that maritime resources were also an aspect of the Chiribaya Baja economy. Additionally, lomas vegetation was only a few kilometers away, at most, and was undoubtedly utilized to some extent. Many of the tombs at this site contained mummy bundles accompanied by minimal, quotidian offerings, and few decorated items were recovered. However, at the central sector of Cemetery 1, tomb construction and grave goods were more elaborate. Overall, the burial pattern of Chiribaya Baja is less ostentatious than at San Gerónimo and Chiribaya Alta, where most individuals are interred with a large number of items. Nonetheless, there is a significant range in status that is apparent within this site.
El Yaral The site of El Yaral (sometimes called "La Yaral" or "Yaral") is located 50 km inland and lies 1000 m above sea level (Rice 1993). The site, which covers about 13 hectares (Lozada & Torres 1991), is located on the terrace and hill slopes above the river (Fig. 6). There are over 300 residential terraces at the site, two identified cemeteries, and two large, communal buildings with stone-foundations (Rice 1993). Radiocarbon dates from domestic contexts place occupation of the site from AD 1027 to AD 1252 (Rice 1993). The mean date for the site is AD 1000 (Tomczak 2001). The vast majority of the domestic architecture at El Yaral is located on terraces that rise above the floodplain. The domestic structures at the site feature cane walls set into wall trenches, and wooden posts. As at the other sites, the structures are generally rectangular and have interior walls that divide the living space into separate rooms. Some adobe lumps have been found on perimeter walls, suggesting that these might have originally been plastered (Rice 1993). The discovery of trenches and wall materials below living floors suggests that the houses were occupied long enough to warrant remodeling. There are two large structures with stone foundations located at the base of the site. Rice has suggested that the larger of these structures served as an area of corporate activity and that the smaller structure was an associated support residence. Lozada and Torres, under the direction of Jane Buikstra, undertook excavations at the two cemeteries located at this site. They report that the burial pattern at El Yaral is considerably different from the burial patterns found in the lower valley sites. While the burials were found in two formal cemetery areas (Buikstra 1995), and the interments were generally of single individuals found in a flexed, seated position and wrapped in textiles, the predominant tomb shape at the site was oval rather than the rectangular form that is commonly found in the lower valley sites. Additionally, the typical burial at El Yaral has a very simple grave assemblage that consists of only a wooden spoon, a gourd or basket, and one or two ceramic vessels. There does not appear to be a range of wealth portrayed in the Yaral burial assemblages as there is at the other Chiribaya sites. The homogeneity of the burial assemblages at Yaral is typical of the preceding mid-valley pattern, where elite did not receive differential burial treatment (Buikstra 1995:253). El Yaral is interpreted as an agro-pastoral site from its location, the items found in middens and grave contexts, and from stable isotope data on human bone from the site (Tomczak 2001). Garcia Márquez (1988) and Rice (1993) report the presence of marine resources in the domestic areas of El Yaral, demonstrating some contact with the coast. The distance between El Yaral and the ocean suggests that marine resources would have had to be preserved in some way (salting or smoking) in order to be transported to El Yaral. This could be an important factor in any potential differences in parasite loads between this site and the lower valley sites. These differences will be further explored and discussed in the following pages. THE PARASITE DATA FROM THE CHIRIBAYA SITES Chagas disease Chagas disease (also America trypanosomiasis) is endemic to Peru and is caused by a hemoflagellate, Trypanosoma (Schizotrypanum) cruzi Chagas, 1909 (Levine 1985). In Peru, its vector is the kissing bug, Triatoma infestans. This disease is characterized by two stages: acute and chronic. The acute phase is characterized by weakness, muscle and bone pain, chills, nervous disorders, heart failure, and anemia (Roberts & Janovy 1996). The anemia associated with Chagas disease is microcytic (Zaidenburg & Segovia 1993) and the marrow is typically hypoplastic (Marcondes et al. 2000). The anemia occurs early in the course of the disease and can become severe (Britton 1969). If the infected individual lives through the acute phase, a chronic phase that is characterized by cardiomyopathy, megaesophagus, and megacolon follows (Nogueira & Rodrigues Coura 1990, Roberts & Janovy 1996). As described by Reinhard and Fink (this volume), megacolon found in prehistoric sites is strong evidence of Chagas disease. One mummy of the 28 observed by Reinhard from Chiribaya Alta exhibited megacolon (Fig. 7). The colon contains an abnormally large accumulation of feces. The fecal pellets are abnormally large and fill the pelvic girdle. This case of megacolon is circumstantial evidence of Chagas disease. Triatomines inhabit human dwellings and hide during the day in crevices, cracks, and thatching. Housing that incorporates vegetation in the walls and roofs provides abundant hiding places for triatomines. The Chiribaya practice of building walls with reed cores would have been especially conducive to triatomine infestation (Fig. 8). In addition, dogs and rodents, including domestic guinea pigs, serve as reservoir hosts for T. cruzi (Nogueira & Rodrigues Coura 1990, Roberts & Janovy 1996). Since the Chiribaya kept guinea pigs and dogs, there was potential for infection.
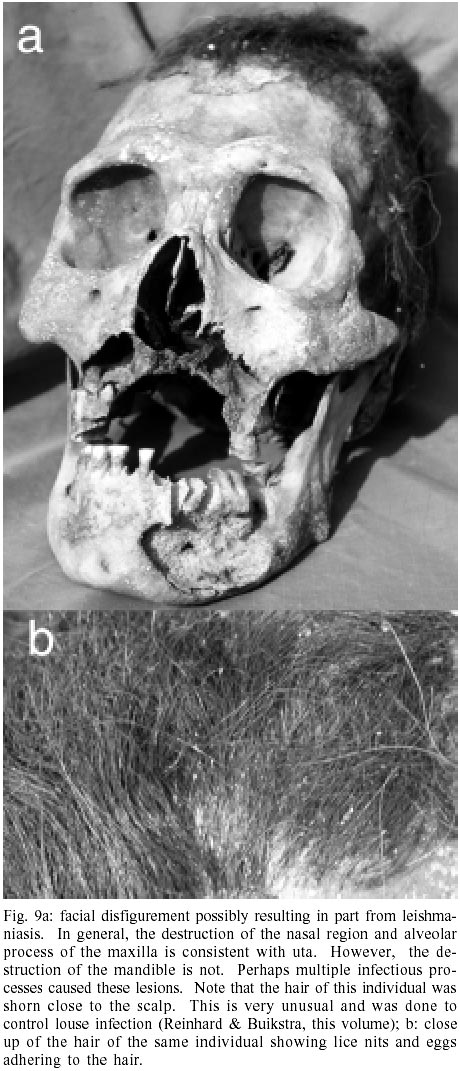
Leishmaniasis Mucocutaneous leishmaniasis, or uta, is endemic to the western slopes of the Andes and is vector borne. Dogs are the most important reservoir for this disease. The vector is the sandfly, probably Lutzomyia noguchii. The sandfly requires high humidity and therefore hides out in crevices or burrows. It is also a poor flier and is nocturnal. Even with these ecological limitations, sandflies are effective vectors of parasitic disease. The parasite that causes uta is classified variably as L. peruviana Velez, 1913 or a variety of L. braziliensis. The parasites are ingested by the sandfly and proliferate in the gut, which is eventually blocked. When the sandfly bites another animal, the parasites are injected into the new host. The parasites metastasize from the primary bite lesion to the mucocutaneous tissues. The infection of the mucocutaneous region results in the destruction of the hard and soft tissues of the nose, palate, and pharynx. Although the destruction of the mucocutaneous tissue can begin before the primary bite lesion is healed, there can be a delay of many years between the initial bite and the mucocutaneous involvement. Of the many diseases caused by Leishmania species around the world, uta is the most disfiguring, and it is clear that the disfigurement impressed ancient Peruvians. The Moche portrayed uta in ceramic art (Wells 1964, Urteaga-Ballon 1991). Indeed, the ancient Peruvian punitive custom of mutilating the nose and lips may have been designed to imitate the disease. Wells warns that the disease could be confused with mycotic infection of the mucocutaneous tissue, based on diagnosis from ceramic representations. Urteaga-Ballon presents a differential diagnosis of the ceramic portrayal. He was able to identify the four stages of the disease from ceramics. The first stage involves a localized ulceration of the nose. The second stage shows soft tissue destruction of both sides of the nose and part of the lip. The third stage shows ulceration of the nose and upper lip. The fourth stage involves the hard tissue, with resorption of the hard palate, nasal septum, and alveolar process, and loss of teeth. Secondary infection can also arise once the mucocutaneous tissue has been destroyed. Individual 317 from Chiribaya Alta exhibits resorption of the nasal aperture, nasal septum, hard palate, and alveolar process, and loss of anterior teeth (Fig. 9). These lytic maxillary changes are consistent with mucocutaneous leishmaniasis. However, the mandible is also effected. A large lytic lesion is present and extends from the anterior alveolar process to a large cavitation on the external surface of the right mandible. It appears that the infection of the mandible initiated at the anterior alveolar margin and then extended to the body of the mandible. Such involvement of the mandible is not described for uta and may represent a secondary infection. Wells' (1964) cautionary note is important in diagnosing uta. He points out that South America blastomycosis and Oroya fever (caused by Bartonella bacilliformis) produce lesions similar to uta. Differential diagnosis in modern patients is dependent on culturing the infective organisms or immunological testing. Unfortunately, with the Chiribaya mummies these techniques were not possible in the last decade. Hopefully, application of PCR genetic testing will be available within the near future to make definitive identification possible. At this point, we present individual 317 as a possible case of uta with the caveat that bacterial or fungal disease agents may have been responsible for his disfigurement.
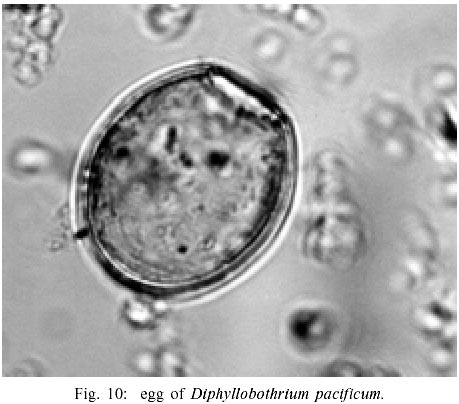
Helminth parasites The most common parasite encountered in Chiribaya materials was Diphyllobothrium pacificum. The diagnosis of D. pacificum is addressed by Reinhard and Urban (this volume). Reinhard's 1991 and 1997 analyses of 28 coprolites recovered from Chiribaya Alta revealed no parasite infections. Reinhard analyzed eight coprolites recovered from San Gerónimo and 11 recovered from Chiribaya Baja. These coprolites were human in origin, but did not come from within mummies. Two of the San Gerónimo coprolites and seven of the Chiribaya Baja coprolites were positive for D. pacificum eggs. In addition, two San Gerónimo coprolites contained trichurid eggs consistent in size with Trichuris trichiura. The egg counts for D. pacificum ranged from 90 eggs per gram of coprolite to 17,800 eggs per gram. The egg counts for T. trichiura ranged above 2,000 eggs per gram. Martinson (2002) also analyzed coprolites and mummies. In her analysis, D. pacificum was, by far, the most common parasite found. Twenty of 29 samples (69%) from Chiribaya Baja, and one of five samples (20%) from Chiribaya Alta were positive for the eggs of this parasite. Coprolites were not available from Yaral, so the helminth parasitism for this site is unknown. CONCLUSION
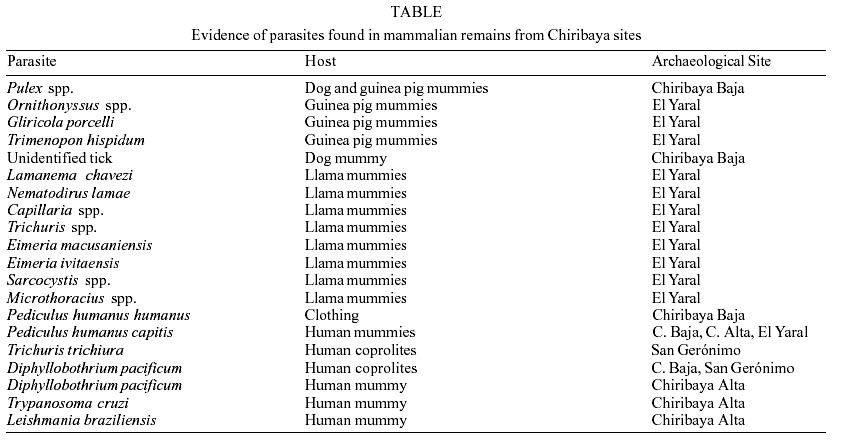
The Chiribaya archaeoparasitology is rich and informative. The combined analyses of human, llama, dog, and guinea pig remains show the diversity of parasite species that lived in prehistoric villages (Table). With regard to guinea pigs alone, four ectoparasite species were found. One of the flea parasites of the guinea pigs and dogs, Pulex spp., also infests humans. With regard to the llamas, seven endoparasite species and one ectoparasite species were found (Leguía & Casas 1999). Among human remains, two types of lice and two helminth parasite species were found. Evidence of megacolon, which is consistent with trypanosomiasis was encountered in a human mummy, and the facial skeleton of one individual exhibited osteolytic lesions that might be related to leishmaniasis. Finally, ticks were encountered on dogs. With regard to human health, fecal-borne helminths rarely infected the Chiribaya. The only fecal-borne helminth found was T. trichiura at San Gerónimo. This makes sense, because San Gerónimo is at the end of the river, downstream from the larger population concentrations at Chiribaya Alta and Chiribaya Baja. In contrast, tapeworm infection occurred at all sites for which we have mummies or coprolites. However, because mummies were found at one site and coprolites at the other sites, the prevalence of infection can not be exactly compared. It appears that D. pacificum infection was rare at Chiribaya Alta. Of 20 mummies analyzed by Reinhard and Martinson, only one contained D. pacificum eggs. In contrast, the numbers of coprolites positive for D. pacificum eggs at San Gerónimo and Chiribaya Baja are quite high. This may mean that there was a greater reliance on fish at San Gerónimo and Chiribaya Baja relative to Chiribaya Alta. However, chemical analysis of some Chiribaya Alta cemeteries revealed high marine food consumption. It may also mean that the fish consumed at Chiribaya Alta was prepared in a way that killed the parasite cysts. In any case, the high numbers of D. pacificum-positive coprolites present at San Gerónimo and Chiribaya Baja is consistent with the archaeological reconstruction of the sites presented above. The focus of these sites on fishing and fish handling for food relates to D. pacificum parasitism. The one possible uta case reveals the nature of cumulative parasitism (Fig. 9a). Whether or not the facial disfigurement is due to a parasitic infection, the louse infestation shows the parasitic consequence of disfiguring disease. As noted by Reinhard and Buikstra (this volume) this individual shows the highest louse infestation intensity of any Chiribaya individual (Fig. 9b). This case suggests that lice opportunistically infested weakened individuals to the maximum benefit of the lice and maximum misery of the host. The parasitism of the dogs and guinea pigs has an obvious influence on human disease. The find of Pulex fleas on these domestic animals shows that the fleas cycled in the Chiribaya villages. As discussed by Dittmar et al. (this volume), the presence of Pulex species has implications for transmission of other non-parasitic diseases such as murine typhus and plague. However, the presence of megacolon, and its implication for trypanosomiasis, in one Chiribaya human mummy raises the importance of dogs and guinea pigs as reservoirs of disease. It is very likely that Chiribaya people kept domesticated guinea pigs in houses, as described for the later Incas, and as is common in the Andes today. The guinea pigs were in the past, and are today, an important source of protein. Dogs and guinea pigs are hosts for T. cruzi. When humans, dogs, and guinea pigs occupy the same houses, the number of available hosts for trypansomiasis increases. Since the construction of Chiribaya houses was ideal for triatomine bug infestation, the association of dogs, humans, and guinea pigs in the same habitation environment raises the potential of transmission to high and perhaps unavoidable levels. Since megacolon is a relatively rare consequence of trypanosomiasis in South America, one observed case is particularly note-worthy in a series of 20 mummies. Certainly, more work based on molecular diagnosis must be done to gain a better idea of the prevalence of this disease among humans and domestic animals. The llamas show a different cycle of diseases than the Chiribaya pastoralists. In contrast to humans, Chiribaya llamas hosted four species of fecal-borne nematodes and two species of fecal born-protozoa. The nematodes are Lamanema chavezi, Nematodirus lamae, Capillaria spp., and Trichuris spp. The protozoa are Eimeria macusa-niensis and Eimeria ivitaensis. The pathology of these combined parasites would have included destruction of the intestinal epithelium, retardation of healing of intestinal lesions, and diarrhea. The fact that these parasites infected llamas suggests that the llamas were frequently herded on moist grazing areas contaminated with eggs and oocysts. The life cycle of Sarcocystis species involves two hosts, a meat-eater such as a dog, and an herbivore. Cysts in the muscle of the herbivore are eaten by the carnivore. The carnivore eventually passes infective oocysts in feces which are then consumed by the herbivore. The cycling of Sarcocystis species in the llamas may have been facilitated by dogs, and implies that dogs might have been associated with llamas. One ectoparasite species of Microthoracius was found. This is a fascinating sucking louse that can reduce wool production and weight gain in infested camelids. Morphologically, the head region of this parasite is well adapted to penetrating the skin and maintaining its place while feeding. Since llamas were a source of wool for the Chiribaya culture, control of Microthoracius spp. infection must have been important. Although much has been accomplished in the archaeoparasitology of the Chiribaya, more can be done with the advent of new techniques (Araújo et al. 1998). The examination for Giardia cysts using immunofluorescence (Gonçalves et al. in press) provides a new approach for the discovery of parasitic protozoa. The development of molecular biology methods for the recovery of T. cruzi ancient DNA (Ferreira et al. 2000) provides the basis for assessing Chiribaya mammalian mummies for Chagas disease. It also opens the possibility of analyzing blood within ancient fleas, lice, and ticks for T. cruzi aDNA ingested from the mammalian hosts. One aspect of future research should include excavations of cane-wall houses for exoskeletons of triatomine bugs. Once collected, the triatomines could be tested for T. cruzi aDNA. Such research would elucidate the ecology of Chagas disease. In addition, a method must be found to definitively diagnose leishmaniasis from ancient hard and soft tissues. With regard to animal parasitism, the coprolites from dog mummies should be examined in the future for Sarcocystis oocysts to determine whether the dogs hosted this parasite. Similarly, coprolites of guinea pigs should be analyzed to determine what endoparasites parasitized these animals. Thus, ten years of archaeoparasitological research of the Chiribaya culture has provided us with considerable insight on the nature of human and animal parasitism in a prehistoric Andean culture. Perhaps more importantly, this work has opened more doors of inquiry that await investigation.
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03029f7.jpg] [oc03029f4.jpg] [oc03029f5.jpg] [oc03029f10.jpg] [oc03029t1.jpg] [oc03029f1.jpg] [oc03029f2.jpg] [oc03029f9.jpg] [oc03029f3.jpg] [oc03029f6.jpg] [oc03029f8.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}