 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 1, January 2003, pp. 129-133 SHORT COMMUNICATION
A Novel Reiterated Family of Transcribed Oligo(A)-terminated, Interspersed DNA Elements in the Genome of Trypanosoma cruzi Newton Valério Verbisck, Márcia Regina Machado dos Santos, David M Engman*, Miguel Angel Chiurillo**, José Luis Ramirez**, Jorge Enrique Araya***, Renato Arruda Mortara, José Franco da Silveira/+
Departamento de Microbiologia, Imunologia
e Parasitologia, Escola Paulista de Medicina, Rua Botucatu 862, 04023-062 São
Paulo, SP, Brasil *Departments of Pathology and Microbiology-Immunology, Northwestern
University Medical School, Chigago, Illinois, USA **Laboratorio de Genetica
Molecular, Instituto de Biologia Experimental, Universidad Central de Venezuela,
Caracas, Venezuela ***Departamento de Tecnologia Médica, Universidad
de Antofagasta, Antofagasta, Chile This work was supported by grants from Fapesp, Capes (Brazil) and Cyted (España) to JFS, Conicit-GR 9900036 (Venezuela) to JLR. NVV was recipient of a doctoral fellowship from CNPq (Brasil). Received 12 August 2002 Code Number: oc03046
We report the molecular characterization of a novel reiterated family of transcribed oligo(A)-terminated, interspersed DNA elements in the genome of Trypanosoma cruzi. Steady-state level of transcripts of this sequence family appeared to be developmentally regulated, since only in the replicative forms the parasite showed expression of related sequences with a major band around 3 kb. The presence of frame shifts or premature stop codons predicts that transcripts are not translated. The sequence family also contains truncated forms of retrotransposons elements that may become potential hot spots for retroelement insertion. Sequences homologous to this family are interspersed at many chromosomes including the subtelomeric regions.
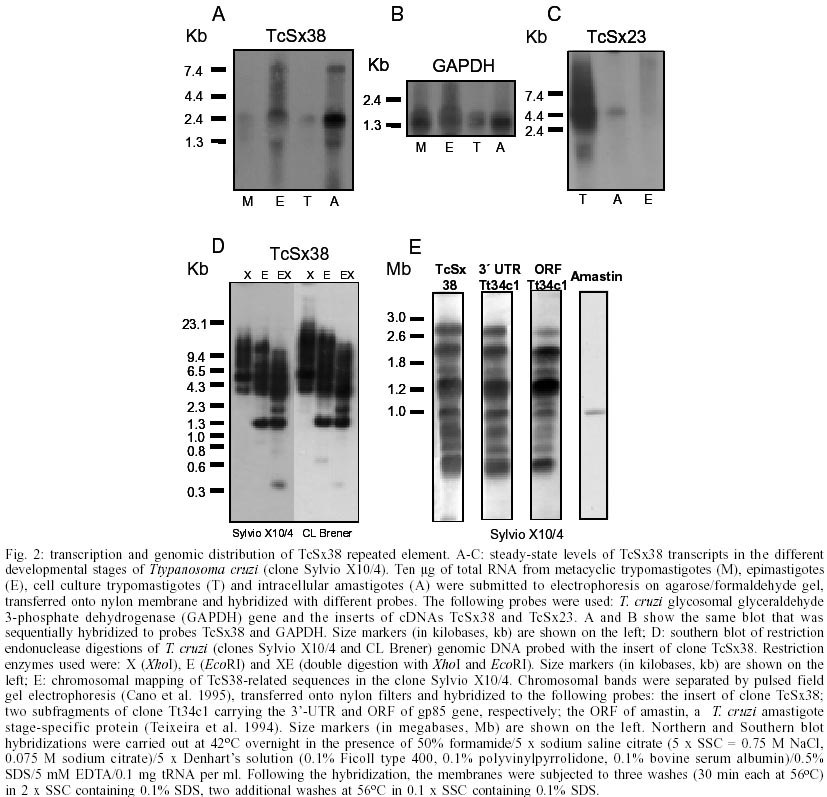
Key words: Trypanosoma cruzi - interspersed repetitive DNA - transcription - retroelement insertion - subtelomeric region The genome of the protozoan parasite Trypanosoma cruzi, the etiological agent of Chagas disease, contains a large number of repeated sequences which may play an important role in genetic rearrangements and control of gene expression (Requena et al. 1996). DNA reassociation kinetic studies have shown that highly and middle repetitive sequences account for nearly 44% of T. cruzi nuclear genome (Castro et al. 1981, Lanar et al. 1981). The repetitious DNA fraction is composed by micro-and mi-nisatellites, short and long interspersed nucleotide elements (SINE and LINE), LTR (long terminal repeat)- and non-LTR retrotransposons (Martin et al.1995, Requena et al. 1996, Araya et al. 1997, Oliveira et al. 1998, Vazquez et al. 1999, 2000, Olivares et al. 2000). Several T. cruzi repetitive elements, such as SINE, LTR- and non-LTR retro-transposons, can be actively transcribed into the poly(A+) RNA fraction (Cotrim et al. 1989, Requena et al. 1994, Martin et al. 1995, Vazquez et al. 1999, 2000, Olivares et al. 2000). The role of these many repeated elements in T. cruzi is still unknown, and it has been speculated that they are involved in shaping and reshaping of the genome causing ectopic rearrangements, modifying and reshuffling existing genes and creating new genes. They can also affect the expression of 5' and 3' adjacent genes by affecting the trans-splicing reaction efficiency (Vazquez et al. 1994, 1999). In this report we characterized a novel sequence family of transcribed oligo(A)-terminated sequences whose members are spread throughout the T. cruzi genome. Although actively transcribed and processed, these sequences had premature stop codons that suggested a lack of translation. In the course of studying genes encoding surface antigens of T. cruzi, a cDNA library constructed in phage l UNI-ZAP XR with poly(A+) mRNA extracted from intracellular amastigotes (clone Sylvio X10/4) was screened with the 32P-labelled insert of a cDNA clone named Tt34c1. Clone Tt34c1 (5488 bp) contains an open reading frame (ORF) for a 85 kDa surface glycoprotein (gp85) and a large 3'-untranslated region (3'-UTR) of 2812 bp but lacks the miniexon and the poly(A) tail (Takle & Cross 1991). According to identity hits in GenBank two types of clones were isolated in these experiments: (a) cDNAs encoding gp85-like glycoproteins (clone TcSx23), and (b) cDNAs sharing similarity with the 3'-UTR of clone Tt34c1 (clones TcSx12, TcSx38 and TcSx42). In this study we further characterized the recombinants TcSx12, TcSx38 and TcSx42. Nucleotide sequences reported in this paper are available in the GenBank database under accession numbers: AF510088 (TcSx12), AF510086 (TcSx38), AF510087 (TcSx42) and AF510037 (TcSx23). Recombinant inserts ranged from 1.5 to 2.1 kb in length, all contained a poly(A) tail and shared considerable identity (~ 92%) at nucleotide level with the 3'-UTR of clone Tt34c1 (Fig. 1A). Sequence comparison among recombinant cDNAs revealed several differences such as the presence of a common ~ 700 bp sequence preceding the poly(A) tail in TcSx38 and TcSx42 but not in TcSx12. In addition, clone TcSx42 has a 157-bp sequence (positions 522 to 679) that is lacking in clone TcSx38. These differences indicated that the three cDNAs originated from different sequence copies. The most striking feature of these transcripts was the presence of multiple stop codons in each frame, indicating that they are not translated. Fig. 1 shows a sequence comparison of cDNAs TcSx12, TcSx38 and TcSx42 with other T. cruzi repetitive sequences reported in the literature. In TcSx38 we identified two subregions (1 and 3) that showed sequence similarity with previously reported T. cruzi repetitive sequences. Subregion 1, from nt 14 to nt 134, has 82.5% and 86% of sequence identity with T. cruzi telomere-associated sequences TcTel9.3 and VATc12, respectively (Chiurillo et al. 1999). Subregion 3, from nt 1570 to nt 2119, presents 50% and 87% of sequence identity with the non-LTR retrotransposons RS13Tc and RS1Tc, respectively (Olivares et al. 2000). In subregion 3 there is also a 41 nt sequence (nt 1592 to 1631) that displays 90% of identity with the 3' end of the E13 repeated element (Requena et al. 1992, Olivares et al. 2000). Subregions 1 and 3 flank a central region of 1434 bp (subregion 2) which is homologous to the 3'-UTR of clone Tt34c1. Clone TcSx12 contains subregions 1 and 2 but lacked subregion 3 which is part of the ~ 700 bp sequence found in TcSx38 and TcSx42, but absent in TcSx12. On the other hand, TcSx42 has subregions 2 and 3 but not subregion 1, this may have been lost during cDNA cloning. It is noteworthy that the 3'-UTR of clone Tt34c1 (Takle & Cross 1991) contains sequences with similarity with the VAT and TcTel telomere-associated sequences but not with non-LTR retro-transposons RS1Tc and RS13Tc. TcSx38 and TcSx42 contain truncated forms of retrotransposons RS1Tc and RS13Tc sequences that have been associated with a hot spot for retroelement insertion in RHS (retrotransposon hot spot) multigene family found in T. cruzi and T. brucei (Bringaud et al. 2002). Steady-state levels of RNAs related to the recombinants were determined by Northern blot analysis using the insert of recombinant TcSx38 as a probe (Fig. 2A). The probe strongly hybridized with a ~ 3 kb transcript and less intensely with two transcripts of around 8 and 1.3 kb in epimastigotes and intracellular amastigotes. A faint hybridization signal was detected with RNAs isolated from bloodstream and metacyclic trypomastigotes. Confirming that TcSx38-related sequences are expressed in the epimastigote forms, a nucleotide sequence identity search of EST (expressed tag sequence) databases with TcSx38 sequence, revealed a high percentage identity with 23 ESTs isolated from a T. cruzi epimastigote cDNA library (Urmenyi et al. 1999). For comparative purposes, when the cDNA TcSx23, which encodes a gp85-like protein, was used as a probe, it hybridized with a ~ 4.5 kb mRNA present in bloodstream trypomastigotes and intracellular amastigotes (Fig. 2C). From these results we concluded that steady-state level of transcripts of TcSx38 is developmentally regulated and differs from that of TcSx23. Taken together, our findings suggested that TcSx12, TcSx38 and TcSx42 belong to a new T. cruzi gene family whose members are transcribed as oligo(A)-terminated sequences and processed to mature RNAs. To further characterize genomic sequences associated to the TcSx38, a Southern blot of genomic DNA digested with several restriction enzymes was probed with the insert of this clone (Fig. 2D). The probe hybridized to multiple genomic bands, suggesting the existence of related sequences arranged in a non tandem manner. A complex hybridization pattern was also obtained for clone CL Brener, the reference clone of T. cruzi Genome Project (Fig. 2D). Chromosomal location of TcSx38 sequences was determined by hybridization with pulsed field gel electrophoresis (PFGE) separated chromosomal bands of clone Sylvio X10/4 (Cano et al. 1995). Implying that copies of TcSx38 are dispersed throughout the genome (rather than concentrated at a single locus), and consistent with the results from the Southern blot analysis, the probe hybridized with eight chromosomal bands of clone Sylvio X10/4 (Fig. 2E) with varying intensities. As an indication that TcSx38-related sequences belong to a multigene family, a nucleotide sequence identity search on GSS (genomic survey sequence) databases with TcSx38 sequence revealed a high percentage identity with 82 GSSs. In fact, according to the equation proposed by Agüero et al. (2000) the copy number of TcSx38-related sequences was estimated to be 73 per haploid genome. For this calculation, we used 47 Mb as the haploid T. cruzi genome, the total number of GSS was 21327, and the size of TcSx38-related sequences was 2119 bp. As described above, clones TcSx38 and TcSx12 also shared sequence similarity with VATc12 and TcTel9.3 sequences located at subtelomeric regions of T. cruzi chromosomes (Chiurillo et al. 1999). To further confirm the presence of TcSx38 sequences at the subtelomeric regions, we have screened a T. cruzi telomeric library constructed in pBelo BAC vector (Chiurillo et al. 2002) with a specific probe of TcSx38 clone (nt 843 to 1767) devoid of VAT or TcTel sequences. BAC telomeric clones (n = 576) were stamped on nylon filters organized in cells of six colonies each and hybridized with TcSx38 probe. Assuming that no bias existed in the cloning and selection procedures, the number of stamped recombinants is slightly higher than a 3-fold representation of each chromosomal end. Positive and negative hybridization controls were included. Thirteen clones strongly hybridized with the probe indicating the presence of TcSx38-related sequences in the subtelomeric regions of T. cruzi chromosomes. The high recombination frequencies at subtelomeric regions may be favorable for the rapid generation of novel variants of this multigene family. In conclusion, our results indicate that TcSx12, TcSx38 and TcSx42 recombinants belong to a new reiterated T. cruzi gene family. They are conserved in sequence, and although their transcripts are polyadenylated, the presence of frame shifts or premature stop codons preclude them from being translated into proteins. Northern blot analysis showed that the steady-state level of transcripts of this gene family is developmentally regulated and mainly expressed in the replicative forms of the parasite (epimastigotes and amastigotes). Members of this multigene family are associated with mobile elements and may be undergoing rapid evolution by recombination and sequence divergence. Our observations suggest that these cDNAs are not derived from read-through transcripts from putative promoters far upstream. Together with other reports of transcribed but not translated sequences, the present work adds more evidences to this puzzling phenomenom that may be a consequence of the all out polycistronic transcription of kinetoplastida, or else it is underlying a more important regulation mechanism for gene expression. The abundance of TcSx38 sequences in transcripts from different T. cruzi developmental forms could suggest that these sequences have a function in the trypanosome genome. Further studies of this sequence family are in progress aimed to unveil the possible function of these elements in the parasite.
ACKNOWLEDGEMENTS
To Dr GAM Cross and Dr SMR Teixeira for kindly providing clones Tt34c1 and amastin, respectively. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03046f2.jpg] [oc03046f1.jpg] |
| |||||||||

{kind=link}
{kind=link}