 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 2, March, 2003, pp. 247-253 Isoenzymatic Variability among Five Anopheles Species Belonging to the Nyssorhynchus and Anopheles Subgenera of the Amazon Region, Brazil Joselita Maria Mendes dos Santos+, Juracy de Freitas Maia, Wanderli Pedro Tadei, Gloria Alícia Diaz Rodriguez
Instituto
Nacional de Pesquisas da Amazônia, Caixa Postal 478, 69011-970 Manaus, AM,
Brasil Research supported by the Program for the Protection of Brazilian Rainforest/MCT, Subprogram C&T/PPD G-7, and CTPETRO. Received
7 May 2002 Code Number: oc03063
An isoenzymatic comparative
analysis of the variability and genetics differentiation among Anopheles species
was done in populations of An. (Nys.) intermedius and An. (Ano.) mattogrossensis
of the Anopheles subgenus, and of An. darlingi, An. albitarsis and
An. triannulatus of the Nyssorhynchus subgenus, with the aim of
detecting differences between both subgenera and of estimating the degree of genetic
inter
Key words: Nyssorhynchus - Anopheles - genetic variability - isozymes - Amazon - Brazil
The main vectors of human malaria in the Amazon are Anopheles mosquitoes of the Nyssorhynchus subgenus. Among them, An. darlingi is the main vector in Brazil, specially in the Amazon, also contributing to malaria endemicity throughout its distribution (WHO 1984, 1988). An. albitarsis sensu lato also transmits malaria in some regions of the country (Deane 1986, Tadei et al. 1993, Tadei & Dutary-Thatcher 2000). This species has recently been found positive by ELISA for Plasmodium falciparum, P. vivax and P. malariae, in the Brazilian states of Amapá and Roraima, and probably plays an important role in malaria transmission in these areas (Póvoa et al. 2001, Silva-Vasconcelos et al. 2002). An. triannulatus sensu lato has been found infected by P. vivax and P. falciparum in immunosorbent (ELISA) and immunoradiometric assays (IRMA) being considered a potential vector (Arruda et al. 1986, Tadei et al. 1988, 1991, Branquinho et al. 1993, Tadei & Dutary-Thatcher 2000). An. darlingi and An. albitarsis belong to the Argyritarsis Section whereas An. triannulatus belongs to the Albimanus Section, both in the Nyssorhynchus subgenus. The Anopheles subgenus has relatively few studies on the biology and genetics of the species comprised in this subgenus, in South America. This, despite the fact that some of the species in the Anopheles subgenus occur sympatrically with malaria vectors of the Nyssorhynchus subgenus, and at times biting man, although they have not been found to be infected with Plasmodium (Tadei et al. 1993). An. (Ano.) intermedius and An. (Ano.) mattogrossensis, of the Laticorn Section, are examples (Harbach 1994). The species studied present a wide geographical distribution. An. darlingi has been recorded from Mexico to North Argentina, from east of the Andes to the Atlantic Coast of South America. In Brazil, it occurs in all states except for Santa Catarina and Rio Grande do Sul (Forattini 1962). An. albitarsis is the species of largest geographical distribution in Brazil, also extending to all countries of South America east of the Andes, from North of Guatemala to the North of Argentina (Rosa-Freitas 1988). An. triannulatus has been recorded throughout South America and in some regions of Central America (Lane 1953, Forattini 1962, Faran & Linthicum 1981). An. intermedius is distributed in all South America up to Mexico in Central America (Forattini 1962, Morales-Ayala 1971). An. mattogrossensis has been registered in all Amazonian countries (Forattini 1962, Garcia & Ronderos 1962, Ferreira 1964). In some regions of their wide distribution, all five species occur sympatrically. An. intermedius and An. mattogrossensis present exophilic habits, whereas An. darlingi, An. albitarsis and An. triannulatus present endophilic and exophilic habits, according to the geographic region (Deane et al. 1988, Tadei et al. 1993). An additional complication for the correct taxonomy and geographic distribution is the fact that adult females of many Anopheles species are difficult to distinguish morphologically and most of them are treated as species complexes. That is the case for the complex An. albitarsis (Kreutzer et al. 1976, Steiner et al. 1982, Rosa-Freitas et al. 1990, Narang et al. 1993). Recent studies with enzymes electrophoresis and RAPD-PCR (random amplified polymorphic DNA-polymerase chain reaction), demonstrated the existence of four species in the albitarsis complex (Rosa-Freitas et al. 1990, Narang et al. 1993, Wilkerson et al. 1995a). Although members of the albitarsis complex have been incriminated as important vectors in the transmission of malaria, their actual role as vectors in a given area might be difficult to determine as they are morphologically indistinguishable (Wilkerson et al. 1995b). Morphological, behavioral and epidemiological differences have been observed in An. triannulatus, rendering more complicated the taxonomic status of this species, which in some cases is considered a highly polymorphic species, but in others two varieties (Consoli & Lourenço-de-Oliveira 1994, Silva do Nascimento 1995). Although in the past the taxonomic status of An. darlingi was controversial (WHO 1984, 1988) due to small morphological variation (e.g. in the scales of hind 3-5 tarsomeres in specimens of the Belize populations, Harbach et al. 1993), little interpopulational variation has been found based on morphological, isoenzymatic and DNA analyses pointing for the existence of a single species (Linthicum 1988, Freitas-Sibajev et al. 1995, Manguin et al. 1999, Santos et al. 1999). Genetic structure studies in Anopheles species have been intensified, mostly in the Nyssorhynchus subgenus, due to its epidemiological importance in malaria transmission. Nonetheless, little is known on the genetics of other species in the Anopheles subgenus in South America, such as An. intermedius and An. mattogrossensis. The present study was conducted to determine the interpopulational genetics divergence and variability among three species of the Nyssorhynchus subgenus, An. darlingi, An. triannulatus and An. albitarsis, which includes the most important species in the transmission of malaria in the New World. It also includes two species in the Anopheles subgenus, An. intermedius and An. mattogrossensis, which have not been incriminated as vectors. MATERIALS AND METHODS
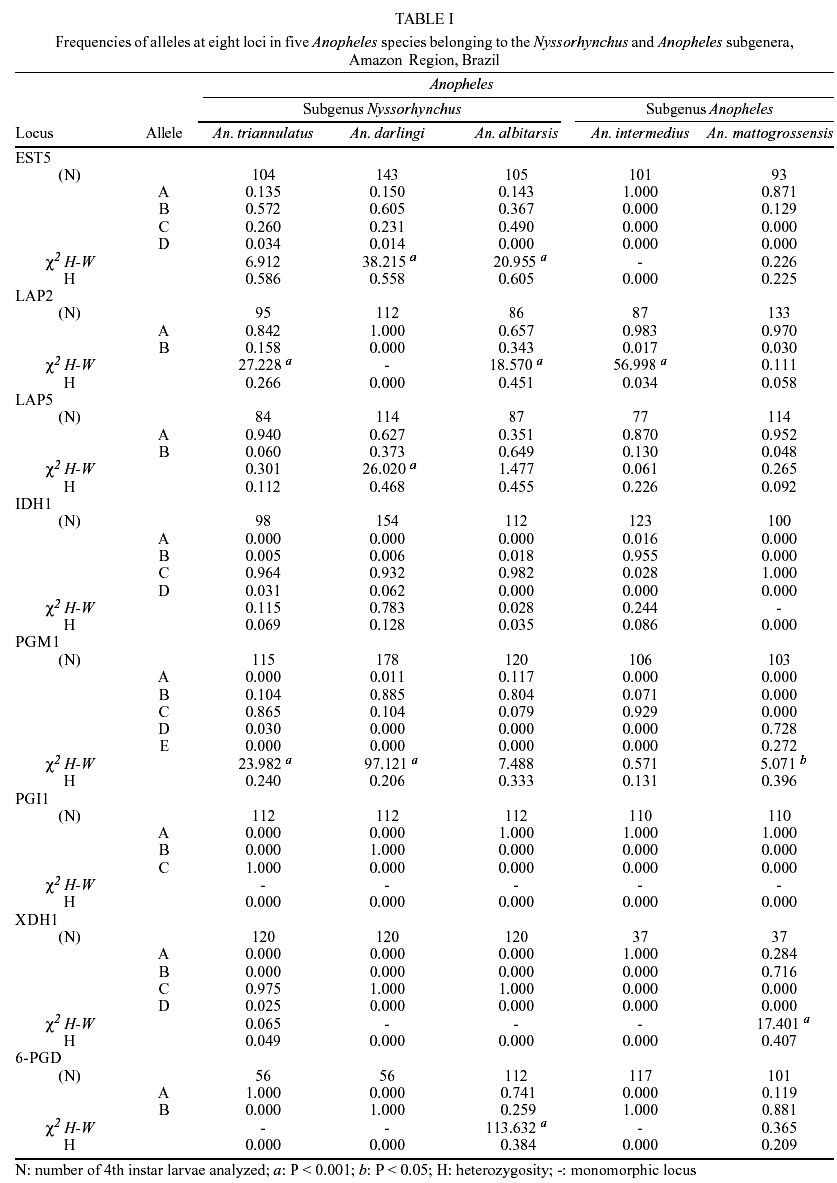
Specimens and collection sites - Specimens of An. darlingi and An. triannulatus were from Mazagão (00º05'15.1''S, 51º17''21.4''W), An. albitarsis and An. intermedius from Rodovia Pacoval (00º02'19.8''N, 51º03'40.7''W), km 4, all near Macapá, State of Amapá. Specimens of An. mattogrossensis were from Janauari Lake (03º08'00.5''S, 60º00'34.7''W), near Manaus. Adults of the subgenus Nyssorhynchus were collected in areas of peri-domiciliary (human and cattle baits) and intradomiciliary (human bait), and those of the subgenus Anopheles were collected only in peri-domiciliary areas (cattle bait) between 18:00 and 22:00 h. Individual females were allowed to lay their eggs separately. Eggs were reared to larvae following the methodology described by Santos et al. (1981). Electrophoretic analyses - Eight loci were analyzed: Esterase (EST5 - E.C.3.1.1.1.), Leucine aminopeptidase (LAP2, LAP5 - E.C.3.4.11), Isocitrate dehydrogenase (IDH1 - E.C.1.1.1.42), Phosphoglucomutase (PGM1 - E.C.2.7.5.1), Phosphoglucose isomerase (PGI1 - E.C.5.3.1.9), Xanthi-ne dehydrogenase (XDH1 - E.C.1.2.1.37) and 6-Phosphogluconate dehydrogenase (6-PGDH - EC 1.1.1.44). Larvae were homogenized individually in 15 µl 0.5% b-mercaptoethanol. Filter paper Whatman no.3 (0.5 cm x 0.6 cm) was used to absorb the homogenate supernatant for horizontal electrophoresis. Isozymes were separeted in two types of electrophoretic support: starch (Sigma) gel, at a concentration of 12% and, starch-agarose gel at concentrations of 2% and 1%, respectively. The three buffer systems used were CA-7: gel buffer-0.009 M Tris, 0.003 M citric acid, pH 7.10; electrode buffer-0.135 M Tris, 0.040 M citric acid, pH 6.90 (Steiner & Joslyn 1979), modificated Poulik: gel buffer- 0.017 M Tris, 0.0023 M citric acid, pH 8.00; electrode buffer- 0.3 M boric acid, pH 8.00 (Contel 1980) and TEMM: electrode buffer-0.1 M Tris, 0.1 M maleic anhydride, 0.01 M EDTA, 0.01 M MgCl2, pH 7.40, and a 1:15 dilution was used in the gel (Harris & Hopkinson 1976) and staining systems were those described in Contel (1980). Statistical analyses - Five larvae from each progeny were used for each single enzymatic system and about 100 individuals, representative of 20 progeny were analyzed to infer the Mendelian inheritance standards of electromorphs for each isoenzyme locus. Isoenzymatic variation among species was analyzed using the Biosys-1 software (Swofford & Selander 1981). Genetic distance was calculated according to Nei (1978). Cluster grouping was done employing the UPGMA method (unweighted pair-group method, arithmetic average) using Biosys-1. RESULTS
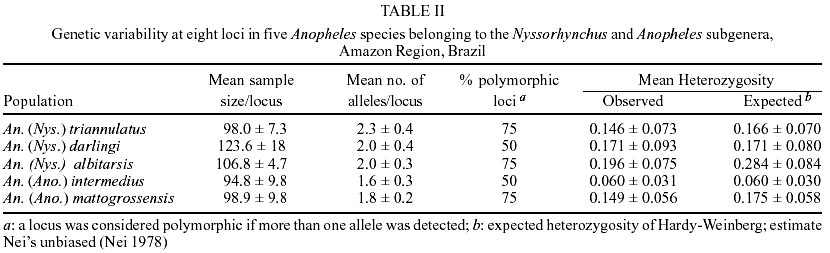
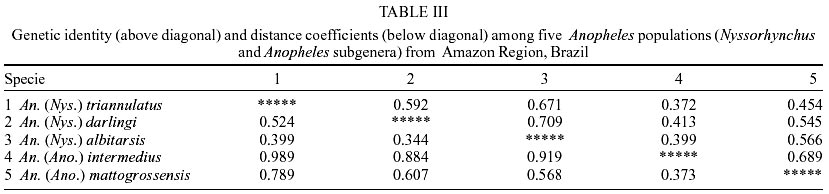
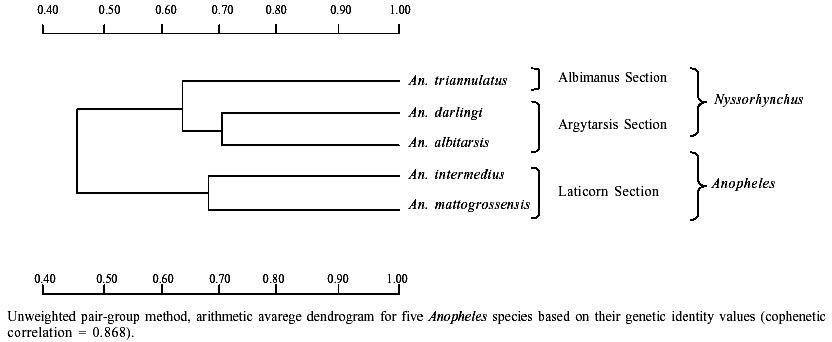
Of the eight loci analyzed, only PGI1 was monomorphic in the five species. The XDH1 locus was polymorphic for An. mattogrossensis and An. triannulatus. The 6PGDH locus was polymorphic for An. albitarsis and An. mattogrossensis. The LAP5 and PGM1 loci were polymorphic for all species. The EST5 locus revealed four alleles in the three species of the Nyssorhynchus subgenus, two alleles for An. mattogrossensis and one allele for An. intermedius, both in the Anopheles subgenus. LAP2 was monomorphic only for An. darlingi and IDH1 only for An. mattogrossensis (Table I). Only IDH1 displayed frequencies agreement to the distribution predicted by random mating for all the species studied. All other loci showed deviation of the genetic equilibrium by Hardy-Weinberg tests. The average number of alleles per loci varied between 1.6 ± 0.30 for An. intermedius to 2.3 ± 0.4 for An. triannulatus. Out of the five species analyzed, An. triannulatus, An. albitarsis and An. mattogrossensis showed larger polymorphism (P = 0.75). An. albitarsis revealed high heterozygozity values (Ho = 0.196 ± 0.075; He = 0.284 ± 0.084). EST5, LAP2 and LAP5 loci (with Ho = 0.605, 0.451, 0.455, respectively) contributing to this result (Tables I, II). Genetic distance (D) values (0.344-0.989) were consistent with interspecific genetic divergence. An. (Nys.) triannulatus and An. (Ano.) intermedius showed higher D (0.989) and An. (Nys.) albitarsis and An. (Nys.) darlingi displayed lower D (0.344), thus separating the subgenera (Table III). The presence of diagnostic alleles in PGI1 (B and C alleles), XDH1 (C and D alleles only in the Nyssorhynchus subgenus, and A and B alleles in the Anopheles subgenus) and PGM1 loci (A allele in the Nyssorhynchus subgenus and to allele E in A. mattogrossensis of Anopheles subgenus) also contributed to these results (Table I). Species were separated in two groups, corresponding to both subgenera according to genetic distance values among species. Group I clustered the three species in the Nyssorhynchus subgenus. An. darlingi and An. albitarsis (Argyritarsis Section) had the highest identity value (I = 0.709) indicating low genetic differentiation and a close phylogenetic relationship, separated from An. tri-annulatus (Albimanus Section). Group II consisted of the two species of the Anopheles subgenus (Figure). DISCUSSION
Genetic, biochemical and morphological studies in anopheline populations have contributed to the elucidation and identification of species complexes, which are of importance to a better knowledge of the malaria transmission and control. Cryptic species are very common in Anopheles. The Nyssorhynchus subgenus, which includes species of great epidemiological importance in Brazil, had studies on genetic differentiation focused mostly on the level of the genetic variability of the populations and for cryptic species recognition (Rosa-Freitas et al. 1990, 1998, Santos et al. 1992, 1999, Narang et al. 1993, Wilkerson et al. 1995a, b, Manguin et al. 1999, Scarpassa et al. 1999). Even so, genetic studies with other sugbenera are still incipient, which might be related to their lack of importance in the malaria transmission. Geographical differentiation in An. darlingi populations has been observed in chromosomes (Kreutzer et al. 1972, Tadei et al. 1982), morphology (Harbach et al. 1993), behavior (Charlwood & Hayes 1978, Charlwood & Wilkes 1979, Roberts et al. 1987, Rosa-Freitas et al. 1992) and hydrocarbon profile (Rosa-Freitas et al. 1992). Isozymes, RAPD and sequencing of the gene ITS2 showed however, no significant differences in Brazilian and other South America populations (Freitas-Sibajev et al. 1995, Manguin et al. 1999, Santos et al. 1999). For isoenzymes, most diagnostic loci that distinguish An. darlingi from other members of the Nyssorhynchus subgenus are monomorphic throughout their distribution area. This was also observed by us for LAP2 and PGI1 loci. PGI1 was also monomorphic for An. albitarsis, An. triannulatus, An. intermedius and An. mattogrossensis for different alleles. The EST5, LAP5, PGM1, IDH1, XDH1 and 6-PGD loci showed variation for two or more species. Our data showed that most loci analyzed were polymorphic for each of the five species, with a relative elevated specific variability (0.50 < P < 0.75) when compared with other studies. Fritz et al. (1995) found a low percentage of polymorphic loci (20.8 < P < 58.3) among populations of An. nuneztovari, An. trinkae and An. rangeli from Brazil, Ecuador, Bolivia and Venezuela. These results might have been influenced by the small sample size, although other variability measures (mean heterozygosity, mean number of alleles) were not affected. In a study based on 33 enzymatic loci (Manguin et al. 1995), low polymorphism was found in populations of An. pseudopunctipennis (from Grenada and Chile, P = 12.1 and 39.4, respectively), An. franciscanus (P = 33.3 from California, USA) and, An. crucians (P = 39.4 from Belize) belonging to the subgenus Anopheles, and An. albimanus (P = 30.3) belonging to the subgenus Nyssorhynchus. According to the authors, the low polymorphism of An. pseudopunctipennis from Grenada is caused by the lack of heterozygotes due to the geographical isolation. However, in the mainland populations of An. pseudo-puctipennis the polymorphism values ranges from 39.4% to 78.8%. Similar results were found by Scarpassa et al. (1999) in a study based on 19 enzymatic loci of the An. nuneztovari populations from Brazil and Colombia, in which the polymorphism varied from 31.3 to 56.6%. Rodriguez (1998) also observed low polymorphism in populations from Manaus of An. (Ano.) intermedius (P = 35.3, Ho = 0.051) and from Amapá of An. (Ano.) mattogrossensis (P = 47.1, Ho = 0.085), based in 18 isoenzymatic loci. In our study, genetic variability (including locus polymorphism, heterozygosity, mean number of alleles/locus) was similar to other species of the Nyssorhynchus subgenus (Narang 1980, Steiner et al. 1982, Santos et al. 1985, 1992, 1999, Santos 1992, Fritz et al. 1995) and of the Anopheles subgenus (Lanzaro et al. 1990, Narang & Seawright 1994). Out of the five species, An. (Ano.) intermedius and An. (Nys.) darlingi displayed the lowest variability and degree of polymorphism for all the eight loci analyzed (P = 0.50). However, regarding heterozygosity, the lowest value found was for An. (Ano.) intermedius (0.060 ± 0.31), whereas An. (Nys.) triannulatus and An. (Nys.) albitarsis showed the highest polymorphism (P = 0.75) and more alleles per locus (2.3 ± 0.4 and 2.0 ± 0.3). Genetic distances were the lowest between An. (Nys.) albitarsis and An. (Nys.) darlingi (0.344), and the highest between An. (Nys.) triannulatus and An. (Ano.) intermedius (0.989). These data are consistent with the interspecific limits for genetic variation, according to the values found for An. rangeli, An. trinkae and An. nuneztovari (0.319-0.440) (Fritz et al. 1995) and for An. pseudo-punctipennis, An. franciscanus, An. crucians and An. albimanus (0.335-2.355) (Manguin et al. 1995). The results from polymorphism presented in this paper showed interspecific genetic divergence separating the species in two groups, in accordance with their subgenera. Considering the genetic distance, the closest genetic relationship belonged to An. darlingi and An. albitarsis while the highest genetic divergence was between An. darlingi and An. triannulatus. Genetic distance values grouped the first two species in the same cluster and separated An. triannulatus from all other species. These data are in part discordant with the phylogenetic relation based on morphologic characters for the subgenus Nyssorhynchus, conducted by Sallum et al. (2000), who grouped An. albitarsis (Argyritarsis Section) and An. triannulatus (Albimanus Section) in the same clade, separated from An. darlingi (Argyritarsis Section).
ACKNOWLEDGEMENTS
To Dr Raul Guerra de Queiroz for help with the English version of the manuscript and the technicians of the Malaria and Dengue Vectors Laboratory for their technical support in the collections and identification of the mosquitoes. To the anonymous reviewers for their comments which greatly improved this manuscript.
REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03063f1.jpg] [oc03063t3.jpg] [oc03063t2.jpg] [oc03063t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}