 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 3, April, 2003, pp. 299-304 SHORT REVIEW Trypanosoma cruzi-elicited CD8+ T Cell-mediated Myocarditis: Chemokine Receptors and Adhesion Molecules as Potential Therapeutic Targets to Control Chronic Inflammation? Joseli Lannes-Vieira Laboratório de Autoimunidade e Imuno-regulação, Departamento
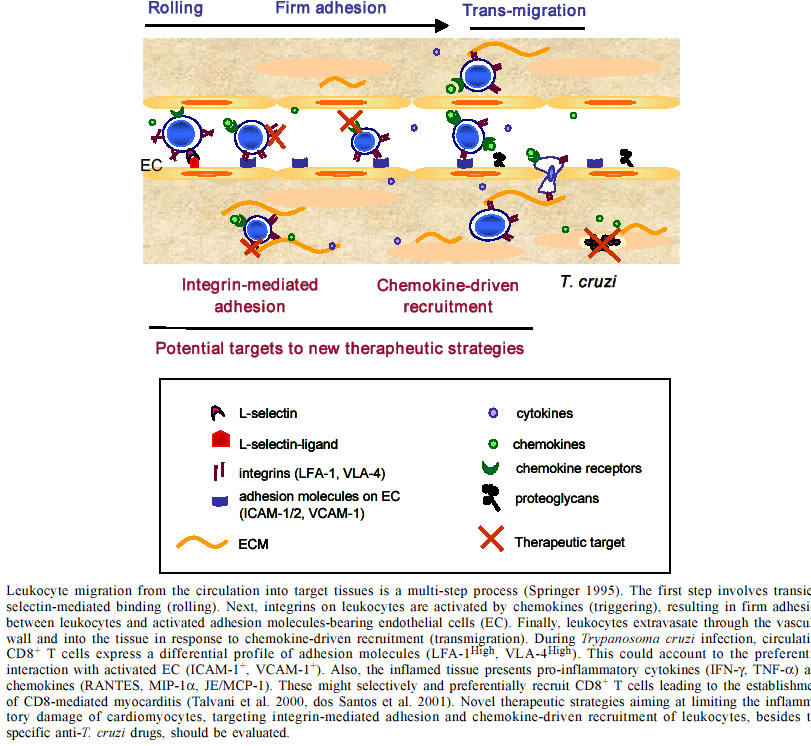
de Imunologia, Instituto Oswaldo Cruz-Fiocruz, This work was supported by grants from Papes-II-Fiocruz, CNPq, Faperj, IOC-Fiocruz, and by a fellowship from CNPq. Received 15 October 2002 Code Number: oc03072 In Chagas disease, during the acute phase, the establishment of inflammatory processes is crucial for Trypanosoma cruzi control in target tissues and for the establishment of host/parasite equilibrium. However, in about 30% of the patients, inflammation becomes progressive, resulting in chronic disease, mainly characterized by myocarditis. Although several hypothesis have been raised to explain the pathogenesis of chagasic myocardiopathy, including the persistence of the parasite and/or participation of autoimmune processes, the molecular mechanisms underlying the establishment of the inflammatory process leading to parasitism control but also contributing to the maintenance of T. cruzi-elicited chronic myocarditis remain unsolved. Trying to shed light on these questions, we have for several years been working with murine models for Chagas disease that reproduce the acute self-resolving meningoencephalitis, the encephalitis resulting of reactivation described in immunodeficient individuals, and several aspects of the acute and chronic myocarditis. In the present review, our results are summarized and discussed under the light of the current literature. Furthermore, rational therapeutic intervention strategies based on integrin-mediated adhesion and chemokine receptor-driven recruitment of leukocytes are proposed to control T. cruzi-elicited unbalanced inflammation. Key words: Trypanosoma cruzi - adhesion molecules - chemokines - inflammation - therapy During an infectious process, the recruitment and migration of leukocytes towards and inside a target tissue is crucial for resolving the infectious and reestablishment of homeostasis. Both recruitment and migration are multi-step processes that depend on the nature and state of activation of the leukocytes, involve pro-inflammatory cytokines, adhesion molecules, and extracellular matrix (ECM) components, being co-ordinated by a mosaic of chemoattractant molecules named chemokines (Shimizu & Shaw 1991, Nathan & Sporn 1991, Del Pozo et al. 1995, Gilat et al. 1996, Sallusto et al. 2000, Gerard & Rollins 2001, Moser & Loestscher 2001). Several studies have demonstrated that CD8+ cells are the predominant cell population in the cardiac tissue of chronic chagasic patients (D'avila Reis et al. 1993, Higuchi et al. 1993, 1997, Reis et al. 1997). Using C3H/HeJ mice infected with the Colombiana strain of T. cruzi we showed the prevalence of CD8+ T cells (CD4:CD8 ratio of 1:3) in the inflamed heart of chronically Trypanosoma cruzi-infected experimental animals. Akin chagasic patients, amastigote forms and parasite antigens were scarce in the chronically inflamed myocardium and associated with the presence of rare IL-4 producing cells, while large amounts of the pro-inflammatory cytokines IFN-γ and TNF-α were detected (Higuchi et al. 1997, Reis et al. 1997, dos Santos et al. 2001). Thus, our results led us to consider that the infection of C3H/HeJ mice with the Colombiana strain of T. cruzi reproduces several aspects of the chagasic chronic myocarditis and is an appropriate model to study the molecular mechanisms involved in the recruitment and accumulation of inflammatory cells into the cardiac tissue leading to the establishment of chronic CD8-mediated T. cruzi-elicited myocarditis. Initially, we showed that the predominance of CD8+ T cells in the cardiac tissue was not a result of the influx of the predominant cell population in the immune compartments, since during the chronic infection CD4+ lymphocytes represent the prevalent T cell population in peripheral blood and lymphoid tissues. Therefore, insight into the mechanisms participating and regulating the selective CD8+ T cell trafficking is crucial when trying to understand how T. cruzi-elicited myocarditis is established. The requirement and functional participation of L-selectin (CD62L), leukocyte function-associated antigen 1 (LFA-1; CD11a,CD18) and very late activation antigen 4 (VLA-4; CD49d,CD29; a4b1) in T cell activation and migration has been extensively demonstrated in other models (Springer 1995, Sprent et al. 1997). For this reason, we investigated the expression of CD62L, LFA-1 and VLA-4 on cells isolated from heart tissue and peripheral blood of T. cruzi-infected mice. A high percentage of circulating CD8+ T cells and most of the CD4+ and CD8+ T cells present in myocardium of T. cruzi-infected animals express an activation phenotype characterized as CD62LLow, LFA-1High and VLA-4High. Interestingly, the distribution of CD4+ and CD8+ T-cells in the myocardium mirrors the frequency of cells expressing the CD62LLowLFA-1HighVLA-4High activation phenotype among CD4+ and CD8+ peripheral blood T cells. In agreement with these results, a recent study demonstrated that patients with a mild form of Chagas disease showed a lower percentage of CD3+a4+ peripheral blood cells, in comparison with those with severe chronic cardiopathy (Laucella et al. 2001). Increased expression of ECM components particularly fibronectin (FN), a VLA-4 ligand, have been previously detected in the myocardium of T. cruzi-infected mice during the acute and chronic phases (Andrade et al. 1989). Studying Colombiana-infected C3H/HeJ mice, we confirmed this finding and showed that a fine interstitial FN mesh present in the inflamed heart involves VLA-4+ CD4+ and CD8+ T cells (dos Santos et al. 2001) as well as the infected and non-infected myocytes (E Roffê & J Lannes-Vieira, unpublished observations). Interestingly, the FN filamentous network present in the inflamed myocardium may function as a pathway in T cell migration process via VLA-4 interactions but may also influence T cell activation. In fact, it was demonstrated that antibodies against VLA-4 increase the anti-CD3-induced T cell adherence to FN and sinergistically induce T cell proliferation (Shimizu & Shaw 1991, Springer 1995). Then, it is reasoned to propose that interactions of VLA-4+CD4+ and VLA-4+CD8+ T cells with the FN mesh present in the inflamed heart could contribute to the perpetuation of the inflammation in this tissue influencing antigen-specific T cell recognition, activation, proliferation, survival and effector activity. Consistently, besides the FN network surrounding VLA-4+ activated mononuclear cells, endothelial cells expressing vascular cell adhesion molecules 1 (VCAM-1), another VLA-4 ligand, were found in the inflamed myocardium, supporting the possibility that VLA-4-mediated interactions might be important for leukocyte entrance in the cardiac tissue, cell migration and positioning towards infected sites as well as persistence of leukocyte activation phenotype inside the inflamed heart (dos Santos et al. 2001). More recently, we have demonstrated that when compared with non-infected or 7 days T. cruzi-infected animals,14 days infected mice present a large proportion of circulating CD8+ T cells expressing the VLA-4+ activation phenotype. Further, the CD8-mediated myocarditis is established on day 28 post-infection. In this moment, most of the mononuclear cells forming focal inflammatory infiltrates surrounding T. cruzi-infected cardiomyocytes are CD8+ lymphocytes, whereas CD4+ cells constituted diffuse infiltrates (AP Marino & J Lannes-Vieira, unpublished observations). Altogether, these results led us to speculate that the predominance of CD8+ T cells in the inflamed myocardium of T. cruzi-infected host might result of the predominance of CD8+ expressing the activated phenotype (CD62LLowLFA-1HighVLA-4High ) in peripheral blood which is established during the early acute infection and persisted during the chronic disease. The use of immunomodulators such as IFN-β able to downregulate the expression of VLA-4 or monoclonal antibodies and antagonists specific for adhesion molecules are being proposed and successfully used to ameliorate inflammatory processes (Silu-Hännien et al. 1995, Yusuf-Makagiansar et al. 2002). Therefore, our results point to the possibility that adhesion molecules may constitute a logic target to control the inflammation elicited by T. cruzi infection. In this context, anti-VLA-4 monoclonal antibodies successfully blocked the ex-vivo adhesion of peripheral blood activated T cells to blood vessels of central nervous system slices obtained from T. cruzi-infected mice. Moreover, anti-VLA-4 antibodies selectively inhibited the migration of activated T cells to the brain tissue of T. cruzi-infected mice (Roffê et al. manuscript in preparation). Presently we are testing the participation of VLA-4-mediated interactions in the establishment of T. cruzi-elicited acute and chronic myocarditis, thus paving the way for the development of a VLA-4-based therapy for T. cruzi-elicited myocarditis. Our previous report showing that a CD8-mediated encephalitis is restricted to the acute phase in the C3H/He mice infected Colombiana strain of T. cruzi (Silva et al. 1999), however, provides evidence that the persistence of chronic myocarditis with the predominance of CD8+ T cells does not only result from the activation phenotype expressed by peripheral blood CD8+ T cells, although it may represent a requirement for the entrance of these cells into the target tissue. Thus, other factors present in the cardiac environment could also be involved in the migration, retention and activation of inflammatory cells leading to perpetuation of the myocarditis in these T. cruzi-infected mice. It is considered that the cytokines produced in the heart tissue during the initial immune response will influence the subsequent immune reaction. In this vein, our data indicate that TNF-α, IFN-γ and IFN-γ-induced chemokines RANTES (regulated upon activation, normal T cell expressed and secreted), MIG (monokine induced by IFN-g) and CRG-2/IP-10 (cytokine response gene 2/interferon g-inducible protein 10), as well as JE/MCP-1 (monocyte chemoattractant protein-1) and MIP1-a (macrophage inflammatory protein 1 a) were found to be the dominant cytokines expressed in situ during acute infection, persisting during the chronic phase of T. cruzi-elicited myocarditis and may contribute to the intense recruitment of activated T cells (dos Santos et al. 2001). Previous report showed that during Listeria mono-cytogenes infection, CD8+ T cells are the main source of MIP-1a and that this chemokine has the ability to preferentially recruit the CD8+ T cell subpopulation (Cook et al. 1999). In addition, in vitro experiments showed that MIP-1a preferentially induces chemotaxis of CD8+ T lymphocytes, whereas MIP-1b is related to the preferential recruitment of CD4+ T cells (Taub et al. 1993). Altogether, these findings led us to propose that IFN-γ and IFN-γ-elicited RANTES, JE/MCP-1 and MIP-1a chemoat-tractants present in the inflamed heart of Colombiana-infected C3H/HeJ mice create a favourable environment for selective and preferential migration of CD8+ T cells towards this tissue, leading to a CD8-mediated myocarditis (dos Santos et al. 2001). The pro-inflammatory cytokines TNF-α and IFN-γ play a major role controlling tissue parasitism during T. cruzi infection (Brener & Gazzinelli 1997, Abrahamsohn 1998). These cytokines are present in the inflamed heart of chronic chagasic patients (D'avila Reis et al. 1993, Reis et al. 1997), and acute and chronic experimentally T. cruzi-infected mice (Powell et al. 1998, Talvani et al. 2000, dos Santos et al. 2001), suggesting that besides contributing to parasitism control they could be also involved in the maintenance of chronic myocarditis (Brener & Gazzinelli 1997, Abrahamsohn 1998). Recently we showed that the heart and central nervous system are the main sites of reactivation of T. cruzi infection in mice lacking functional genes for IFN-γ and IL-12, respectively (Michailowsky et al. 2001). This reactivation was characterized by intense inflammation accompanied by remarkable enhancement in parasitism, particularly resembling the parasite-elicited mass encephalitis often found in chronic chagasic patients with AIDS (Rocha et al. 1994). Importantly, during reactivation of infection the inflammatory processes in the central nervous system from IL-12 knockout mice were mostly devoid of CD8+ T cells and mainly composed of polymorphic nuclear cells, macrophages and CD4+ T cells, whereas in the heart CD8+ T cells predominate over CD4+ T cells. Consistent with the hypothesis that CD8+ T lymphocytes are the major source of IFN-γ and IFN-γ-inducible chemokines that orchestrate the preferential CD8 migration during T. cruzi infection, we found IFN-γ producing cells in the heart, but not in the central nervous system of these animals (V Michailowsky & J Lannes-Vieira, unpublished observations). Thus, considering the importance of CD8+ T cells in resistance to T. cruzi (Tarleton 1990, Tarleton et al. 1994), the increased susceptibility to parasite replication and mass encephalitis could be explained by the lack of CD8 T cell migration and local production of IFN-γ and IFN-γ-inducible chemokines in IL-12 deficient mice. Our results also showed that in contrast to IFN-γ knockout mice, splenocytes from IL-12 knockout mice infected with T. cruzi produced low levels of IFN-γ upon stimulation with parasite antigens. Thus, our data suggest that the level of IFN-γ deficiency is a major determinant of the site of reactivation of T. cruzi infection in immunocompromised host (Michailowsky et al. 2001). Chemokines are small (8-14 kDa) inducible cytokines that recognize a large group of 7 transmembrane-spanning G-protein-coupled serpentine receptors displayed on the leukocyte surface and are involved in normal trafficking of leukocytes to both lymphoid and nonlymphoid organs and recruitment of these cells to sites of injury and infection. Moreover, chemokines also appear to play an important role in immune regulation, mediating leukocyte activation, costimulation and differentiation during innate and adaptive immune responses (Sallusto et al. 2000, Gerard & Rollins 2001, Moser & Loestscher 2001). Recent studies have shown that human macrophages in vitro infected with T. cruzi (Villalta et al. 1998), and in vivo and in vitro T. cruzi-infected peritoneal macrophages and cardiomyocytes produce RANTES, MIP-1a and JE/MCP-1 and respond to these chemokines increasing T. cruzi uptake, enhancing nitric oxide production and controlling parasite replication (Aliberti et al. 1999, Machado et al. 2000). Further, using knockout mice infected with T. cruzi we showed that both IFN-γ and TNF-α are essential for the production of RANTES and MIP-1a, respectively (Aliberti et al. 2001). Also, IFN-γ enhanced the in vitro production of RANTES and IP-10 by macrophages infected with T. cruzi or treated with glycosylphosphatidy-linositol-anchored mucin-like glycoproteins of try-pomastigotes (tGPI-mucins) (Talvani et al. 2000, Aliberti et al. 2001). Thus, it is possible that IFN-γ-induced chemokines present in the cardiac tissue o T. cruzi-infected individuals could also participate in parasitism control. Considering a general model for Chagas' pathogenesis, the results discussed above suggest that the predominance of CD8+ T cells in the myocardium of T. cruzi-infected individuals reflects the profile of adhesion molecules and chemokine receptors displayed by the circulating CD8+ T cells and point to the possibility that multiple IFN-γ-inducible molecules present in the inflamed tissue contribute to the genesis and maintenance of T. cruzi-induced myocarditis. The corollarium of this hypothesis is that activated CD8+ T cells present in the inflamed heart as well as a large proportion of circulating CD8 lymphocytes of T. cruzi-infected individuals differentially bear CC chemokines receptors such as CCR1, CCR3, CCR5 (Proudfoot 2002), in comparison with CD4+ T cells. Interestingly, a differential expression of CCR5 was observed on CD3+CD8+ peripheral blood T cells obtained from patients with cardiomyopathy compared with indeterminate patients or uninfected individuals (Gomes et al., manuscript in preparation). In agreement with this finding, it has been shown that a CCR5 promoter point mutation (CCR5D32) was significantly increased in asymptomatic patients compared with patients with cardiomyopathy (Calzada et al. 2001). Moreover, it has been reported that most CD8+ T cells present in the heart of T. cruzi-infected mice express CCR5 in heart (Teixeira et al. 2002, Marino et al., manuscript in preparation). Altogether, these results and our data discussed above led us to consider that CC chemokines, specially RANTES and MIP-1a, and CC receptors, mainly CCR5, could be involved in differential cell migration and pathogenesis of T. cruzi-elicited CD8-mediated myocarditis. To test this possibility we used Met-RANTES, a N-terminally modified human RANTES that inhibits agonist activity at CCR1 and CCR5 (Proudfoot et al. 1999), to modulate the establishment of T. cruzi-elicited acute myocarditis. Our results showed that Met-RANTES treatment increased the survival of T. cruzi-infected animals when compared to saline treatment. Moreover, Met-RANTES treatment significantly decreased the inflammatory infiltrates due to CD4 and CD8 T cells, but decreases or does not interfere, depending on the therapeutic scheme used, with the heart parasitism (Marino et al., manuscript in preparation). These results indicate that the massive influx of CC chemokine receptors-bearing inflammatory cells into the heart tissue is not crucial for cell-mediated anti-T. cruzi immunity, however it seems to be critical for T. cruzi-elicited immunopathology. Thus, the beneficial role of Met-RANTES may relay on the reestablishment of balanced inflammation, however further investigation are required to unravel the beneficial effector mechanisms of Met-RANTES during T. cruzi infection. Anyhow, the inhibition of CC chemokine receptors might become an attractive therapeutic strategy for further evaluation during T. cruzi infection. Thus, further studies may provide additional insights into the mechanisms by which adhesion molecules, chemokines and chemokine receptors contribute to the establishment and regulation of the immune response in the inflamed heart during T. cruzi infection and may unravel novel therapeutic strategies aimed at limiting the inflammatory damage to cardiomyocytes, targeting integrin-mediated adhesion and chemokine-driven recruitment of leukocytes besides specific anti-parasite drugs (Figure). ACKNOWLWDGEMENTS To the MSc and PhD students of the Laboratório de Autoimunidade e Imuno-regulação (Departamento de Imuno-logia, IOC-Fiocruz) and collaborators that contributed to the work discussed here. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03072f1.jpg] |
| |||||||||

{kind=link}