 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
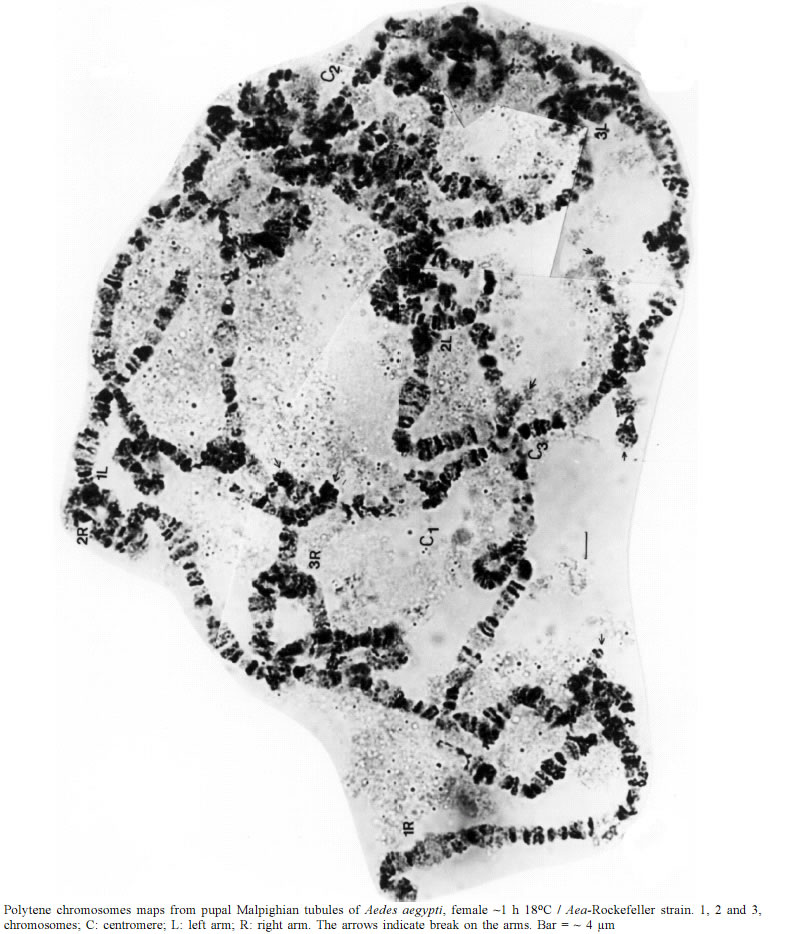
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 3, April, 2003, pp. 387-390 SHORT COMMUNICATION A Technique for Preparing Polytene Chromosomes from Aedes aegypti (Diptera, Culicinae) Jairo Campos/+, Carlos Fernando S Andrade, Shirlei M Recco-Pimentel* Departamento de Zoologia *Departamento de Biologia Celular, Instituto de Biologia,
Universidade Estadual de Campinas, 13084-971 Campinas, SP, Brasil This research was supported by Capes. Received 26 August 2002 Code Number: oc03087 Polytene chromosome preparations were obtained from larval, pupal and adult female Malpighian tubules of Aedes aegypti. The Malpighian tubules of the pupae (0-4 h old) from larvae reared at 20oC provided the best cytogenetic analysis. The interaction of nucleic acids and proteins that influence the spreading of the chromosomes could be reduced with the preparation technique of the sheets submitted to a stronger treatment starting with the hypotony of tissue and successive bathings with acetic acid. A simple technique should facilitate molecular cytogenetics used in the location of resistance and vector competence genes. Key words: cytology - mosquito - vector Studies of the cytogenetic and molecular biology of Anophelinae species can be performed by the analysis of polytene chromosomes structure. Preparation of polytene chromosomes in Culicinae species is difficult and the available techniques are not always reproducible. Although such analyses remained refractory for some species of mosquitoes (e.g. Aedes aegypti), Malpighian tubule polytene chromosomes are an excellent material for detailed approaches in the cytogenetic analysis of Culex quinquefasciatus (Campos 2002). In the present study, polytene chromosome slides were obtained from pupal Malpighian tubules of A. aegypti and compared with published data. Polytene chromosome preparations were obtained using larval, pupal and adult female Malpighian tubules of A. aegypti. The individuals were reared under standard conditions (20 ± 2oC, 70 ± 10% RU). The larvae were fed ad libidum with yeast. Abdomens of larvae, pupae or adults were dissected in Ringer's solution and the Malpighian tubules transferred to a siliconized coverglass with distilled water at 3oC for 1-2 min, then removed and placed in a drop of modified Carnoy's fixative (3:1 95% ethanol: acetic acid) for 1 to 3 min and 60-100% acetic acid added for 2 to 4 min, subsequently stained with 1% aceto-orcein for 4-5 min. The Malphigian tubule cells were dissected in lactoacetic acid (85% lactic acid-100% acetic acid, 0.55: 0.45) or lactic acid 80%; all cytoplasmatic components were removed and the chromosomes were left for a minimum of 20 to 48 h at 3oC. Finally 60-100% acetic acid was also added. Squashing was effected by tapping gently and patiently to spread the chromosomes. Several bathings of 60% acetic acid in the slide allowed good spreading of the chromosomes with complete analysis of the banding pattern. The Malpighian tubules of the pupae (0-4 h old) from larvae reared at 20oC provided the best cytogenetic analysis. White, gray or creme pupae with a transparent thorax (< 30min) are the best material. Conspicuous chromosomal banding pattern, amorphous regions and puffs characterized the pupal Malpighian tubule polytene chromosomes of A. aegypti (Figure). The pupal chromosomes, when compared with larval (salivary glands) chromosomes (Sharma et al. 1978), show certain technical advantages: (1) pupal Malpighian tubules are very easy to dissect in comparison with salivary glands of larvae and (2) higher band resolution in the pupal chromosomes is obtained. The polytene chromosomes obtained in the pupal Malpighian tubules showed fragile structures (Figure), however the availability of suitable slides was 4% (58 out of the approximately 1,383 slides). Chromosome polytene physical maps aimed at correlating with genetic linkage maps could be developed, relying on fluorescence in situ hybridization (FISH) techniques. This is being done for A. aegypti with the meta-phase chromosomes (Brown & Knudson 1997, Brown et al. 1995, 2001). In this way and based on the approaches already effected with insecticide resistance probes, research on vector competence and the location of the any cytogenetic markers should be explored. Polytene chromosomes provide a distinct advantage in generating and integrating genetic and physical maps (Severson et al. 2001). The technical difficulties in the preparation of polytene chromosomes of Aedes genus are evident (Sharma et al. 1978, 1986) and are reflected in the lack of papers dealing with this material. Several problems have been already suggested as causal of the low quality of polytene chromosomes preparations which could be suitable for analysis in Aedes and Culex: (1) Sutton (1942) suggested the presence of weak points, which can be assumed now as being heterochromatic areas where the chromosomes break easily (Semeshin et al. 2001); (2) the great length of chromosome arms (Kitzmiller 1963) should influence chromatic interactions; (3) the inter- and intra-chromosomal connections, or ectopic pairing (French et al. 1962, Verma et al. 1987) resulting from regions of highly repetitive DNA (Rai & Black IV 1999, Severson et al. 2001); (4) surface adhesions (Rai 1967 apud Sharma et al. 1978) that have been observed in Anopheles funestus and is dependent on b-heterochromatin (Sharakhov et al. 2001) and (5) asynapsis observed in the polytene complement (Zambetaki et al. 1998). The larvae reared in low temperature, 18-20oC (Kanda 1970, Sharma et al. 1978) and the larvae's physiological characteristics (Verma et al. 1987) can improve the chromosome spread and the quality of the salivary gland chromosome preparations. This was verified here with Malpighi tubule chromosomes of A. aegypti. Additional observations in preparations of Malpighi tubule chromosomes of A. albopictus and Ochlerotatus fluviatilis allowed the verification that the genoma size and the polyteny degree can influence the quality of the preparations. For pupae of A. albopictus, the preparations were of low quality, very inferior to those of A. aegypti, while for larva of O. fluviatilis the preparation presented good polyteny with clear resolution of bands and well-spread chromosomes, superior to those found for A. aegypti. It is known that Brazilian populations of A. albopictus present a larger genome (Kumar & Rai 1990) than that of A. aegypti and another species of Aedes (Rao & Rai 1987, Knudson et al. 1996). This and the low polyteny observed have surely determined the inferior quality of the A. albopictus preparations that present a smaller polytene nucleus in Malpighi tubules and poorly spread chromosomes. On the other hand in O. fluviatilis, the degree of polyteny observed was larger than that registered in this work for A. aegypti. It has been suggested that the amount of heterochromatin and its distribution in the chromosomes are the cause for lack of band resolution in the polytene chromosome preparations and that it restricts their spreading (Knudson et al. 1996, Rai & Black IV 1999). But the results presented here are not in agreement with this idea. It can be assumed that, rather than a direct involvement of the heterochromatin, the determinating factor for good polytene chromosome preparations from Culicinae has to do more with the polyteny degree, the physiologic state and the techniques used. The first can be particular for the strain, therefore genetically determined, as it was observed for C. quinquefasciatus strains (Campos 2002). The influence of the physiologic state on the spread is not only related to the polyteny degree but also to the development in favorable environmental conditions (low larval density for volume and surface of the medium, feeding and temperature), can determine that RNA and specific nuclear proteins have differential expression facilitating the chromosome spread. The interaction of nucleic acids and proteins, that also influence the chromosome spread, could be reduced in the technique preparation of the slides by a stronger treatment starting with the hypotony of tissue and successive bathings with acetic acid. Acid treatment helps for well spread chromosomes of C. quinquefasciatus (Achary 1994) and this can be associated with acid proteins that are easily extracted with the treatment. In the case of the lack of spreading in A. aegypti, this can have to do mostly with non-acid proteins (non-histones). From the molecular view point, based in the above results, it can be affirmed that more than the amount of heterochromatin, the genome size, the interspersion pattern (repetitive DNA/single DNA) and the protein composition are factors that influence the spreading of chromosomes. C. quinquefasciatus, has an intermediate genome size and an intermediate to short- or long-interspersion pattern, while A. aegypti possesses a larger genome with a short interspersion pattern (Severson et al. 2001). The percentage of repetitive DNA of C. quinquefasciatus is larger than in A. aegypti, 80% against about 60% (Warren & Crampton 1991, Knudson et al. 1996, Brown et al. 2001). Thus, the statement of Severson et al. (2001) that the problem of lack of polytene chromosome spreading is caused by highly ectopic pairing, resulting from areas of highly repetitive DNA, is at least, partly unsustainable because of the observation of better spreading in C. quinquefasciatus than in A. aegypti. Conservation of chromosome arms among higher taxa is relatively common in Diptera. Comparative linkage maps for the mosquitoes, C. pipiens and A. aegypti, indicated that the chromosome 1 is highly conserved between the two species and several homologous loci exist among the arms of the chromosomes 2 and 3 (Mori et al. 1999). Starting with the present work, a comparison will be made between the chromosome maps of A. aegypti and C. quinquefasciatus establishing analogies with those of the linkage maps. In this way, the association of arms can be evaluated among these species by means of the existence of homologies of chromosome landmarks and band groups. The pupal Malpighian tubule polytene chromosomes showed conspicuous structural characteristics suitable for their use in the location of resistance and vector competence genes. The chromosome maps are fundamental tools to provide good cytogenetic analyses of this mosquito, which is of medical and economic importance. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03087f1.jpg] |
| |||||||||

{kind=link}