 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 4, June, 2003, pp. 435-442 Trypanosoma (Herpetosoma) rangeli Tejera, 1920 - An Updated Review Felipe Guhl/+, Gustavo Adolfo Vallejo* Centro de Investigaciones en Microbiología
y Parasitología Tropical, Universidad de los Andes, A.A. 4976, Bogotá,
Colombia *Laboratorio de Investigaciones en Parasitología Tropical,
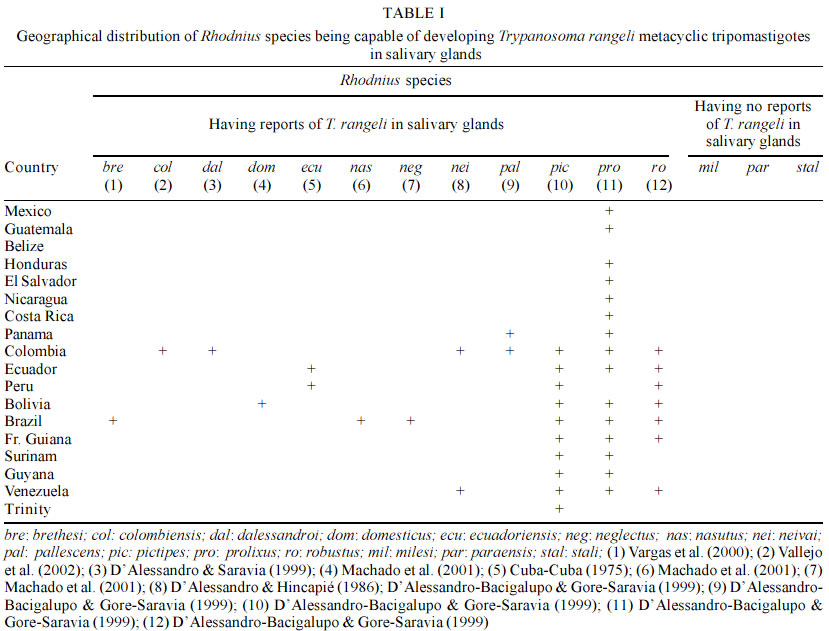
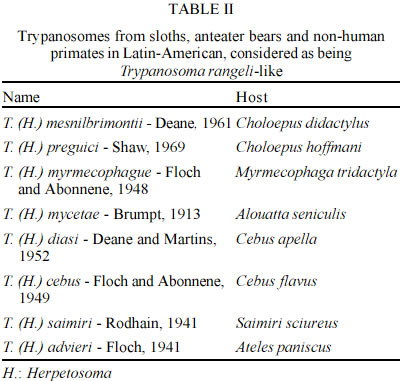
Departamento de Biología, Universidad del Tolima, Ibagué, Colombia Received 25 March 2003 Code Number: oc03097 Trypanosoma rangeli, a parasite generally considered non-pathogenic for man, is the second species of human trypanosome to be reported from the New World. The geographical distribution of T. rangeli often overlaps with that of T. cruzi, the same vertebrate and invertebrate hosts being infected. Their differentiation thus becomes of real, practical importance, particularly as they share approximately half the antigenic determinants recognized by the humoral response. Little is known about the life cycle of T. rangeli in the vertebrate host, although thousands of human and wild animal infections have been reported. Recent studies have revealed 2 major phylogenetic lineages in T. rangeli having different characteristics, thus leading to better understanding of the epidemiology and interactions with this parasite's vertebrate hosts and triatomine vectors. Based on further genetic characterization analysis, the authors have proposed 2 alternative hypotheses and consider that T. rangeli could have had clonal evolution or have been subjected to speciation processes. Key words: Trypanosoma rangeli - vectors - reservoirs - biochemistry - genetic characterization Trypanosoma rangeli is the second trypanosome species frequently found to be infecting humans in Latin-American countries. Even though T. rangeli is apathogenic for the vertebrate host, its biological and epidemiological characteristics are studied within the context of the biology and epidemiology of T. cruzi (the causative agent for Chagas disease), as in some areas of America both species produce mixed infections in vectors making species' morphological differentiation difficult (Vallejo et al. 1988) or producing mixed infections in vertebrates, generating crossed serological reactions complicating the specific diagnosis of Chagas' infection (Guhl & Marinkelle 1982, Guhl et al. 1985, 1987). This parasite thus generates important practical consequences in the epidemiology of American trypanosomiasis, as the presence of T. rangeli in those areas which are endemic for T. cruzi poses the need for devolving methodologies guaranteeing the correct diagnosis of Chagas disease. THE PARASITE T. rangeli is a trypanosome of man and other vertebrates, characterized by possessing a similar morphological structure to that of T. lewisi, the species belonging to the Herpetosoma subgenus that are transmitted by reduviid bite. The sanguineous form of T. rangeli is slender, being 26 to 34 µm in length, including the free flagellum. It has a small, round, subterminal kinetoplast; the nucleus is close to the middle of the body and the undulating membrane is well-developed. Triatomid vectors become infected on ingesting animals' blood with T. rangeli. The parasite subsequently develops within the insect's intestine, producing forms similar to amastigotes, short epimastigotes, large epimastigotes spheromastigotes and tripomastigotes (Vallejo et al. 1988). The most outstanding biological characteristics (regarding T. rangeli in a vector) are presented by flagellate penetration from the intestine towards the haemolymph and salivary gland invasion accompanied by metacyclic tripomastigote formation becoming infectious for the vertebrate host through an insect's bite. A trypanosome must show morphology in a vertebrate's blood compatible with the Herpetosoma subgenus' characteristics to become considered sensu stricto as being T. rangeli and it must be able to infect triatomine vectors with concomitant infectious metacyclic tripomastigote formation in the salivary glands. Therefore, according to the foregoing, flagellates having Herpetosoma subgenus morphology isolated from triatomid intestine or vertebrate blood, capable of developing within the intestine, but having been biologically proved as being incapable of invading the haemolymph and producing metacyclic tripomastigotes in triatomid vector salivary glands, cannot be considered to be T. rangeli, thus being defined as being similar to T. rangeli or T. rangeli-like. BIOLOGICALLY PROVEN T. RANGELI VECTORS Triatomine insects represent the biologically proven vectors for T. rangeli, being capable of developing infectious metacyclic tripomastigotes in their salivary glands in natural or experimental conditions. The development of T. rangeli in salivary glands and its vectorial capacity through biting in natural or experimental conditions has been proved in 12 out of the 15 species of Rhodnius currently described (Table I). In addition to the aforementioned vectors, natural T. rangeli infection has only been reported in the salivary glands of T. dimidiata captured in human dwellings in Colombia, being transmitted to a vertebrate host via bite (Marinkelle 1968a); recent studies in our laboratory have shown that experimental intrafemoral infection with T. rangeli forms in T. dimidiata infect the salivary glands after 40 days in 4% of the studied insects (unpublished data). The simple presence of flagellates similar to those of T. rangeli in an insect's intestine does not constitute a test of the insect's vectorial capacity, as the definitive evidence consists of demonstrating the presence of metacyclic tripomastigote in salivary glands being infective for a vertebrate. T. RANGELI-LIKE TRYPANOSOMES BEING RECORDED IN OTHER TRIATOMINES T. rangeli-like trypanosome have been observed in the intestine (but not in the haemolymph or salivary glands) of 16 species corresponding to 5 triatomid genera (Cavernicola, Eratyrus, Dipetalogaster, Panstrongylus and Triatoma), constituting a conclusive test that they are not T. rangeli vectors (D'Alessandro 1976, D'Alessandro-Bacigalupo & GoreSaravia 1992, 1999). REPORTS OF T. RANGELI-LIKE TRYPANOSOME IN ANIMAL BLOOD Several species of trypanosome belonging to the Herpetosoma subgenus have been named and reported in sloths, anteater bear and non-human primates in several Latin-American countries (Table II). Due to these parasites being developed within the intestine but not in the haemolymph or salivary glands of triatomid vectors, different authors have considered this species as being T. rangeli-like (Hoare 1972, D'Alessandro 1976). RESERVOIRS Herpetosoma subgenus species usually have specific hosts; however, T. rangeli presents a wide range of vertebrate reservoirs distributed among 5 orders: Edentata, Marsupialia, Carnivora, Rodentia and Primates (D'Alessandro 1976, D'Alessandro-Bacigalupo & Gore-Saravia, 1992, 1999, Cuba Cuba 1998). Different species, including rats, mice, hamsters, rabbits, dogs, guinea-pigs, bat and primates have all been experimentally infected with T. rangeli. HUMAN INFECTION In spite of human infections by T. rangeli being frequently registered in different American countries, it is difficult to establish a global figure due to serologically crossed reactions with T. cruzi (Guhl & Marinkelle 1982, Guhl et al. 1985, 1987). However, several publications have demonstrated the presence of T. rangeli in humans by direct examination, haemoculture or xenodiagnosis leading to more than 2,600 cases of human infection being registered: 1,144 cases in Venezuela; 1,117 in Guatemala; 181 in Panama; 121 in Colombia; 61 in El Salvador; 4 in Costa Rica; and 2 cases in Peru (D'Alessandro 1976, D'Alessandro-Bucigalupo & Gore-Saravia 1992, Grisard et al. 1999). Coura et al. reported the first case of human infection by T. rangeli in Brazil (1996). BIOLOGICAL CHARACTERISTICS Biological characterization of T. rangeli has shown the existence of intra-specific variations. For example, the experimental transmission of T. rangeli to a vertebrate host by means of infected triatomid faeces was published by Coutinho and Nussennzweig (1952). Similar data was obtained by HerbigSandreuter (1955), who showed infection in 2 out of 20 lactating mice, and Grewal (1956) who described this parasite's transmission by the same route. On the other hand, other authors (Tobie 1964, Cuba-Cuba et al. 1972, D'Alessandro 1976, Hecker et al. 1990, Steindel et al. 1991) have not been able to link infection in mice with inoculating flagellates present in the faeces of T. rangeli infected triatomids. In addition to the main natural T. rangeli infection route (which is the inoculative route via an insect's saliva), experimental infection of mammals by inoculating forms of T. rangeli culture from different strains has been described (D'Alessandro 1976, Steindel et al. 1991). The literature has shown heterogeneous results regarding T. rangeli persistence in vertebrate host blood. In this sense, Añez (1981) and Añez et al. (1985) could not prove the parasite's presence by xenodiagnosis or haemoculture in mice and Didelphis marsupialis following 50 or 60 days' infection with a T. rangeli strain originating from Venezuela. On the other hand, Steindel (1993) has shown the presence of T. rangeli by haemoculture in mice following a period of 7 months' infection. The parasite has been detected after 13 months' (Paredes & Paredes 1949) and after 18 months' infection (Groot et al. 1950, Groot 1954) in natural and experimental human infection using the same technique. VECTOR SUSCEPTIBILITY TO INFECTION WITH T. RANGELI It has been observed that the triatomids (particularly from the Rhodnius genus) are susceptible to T. rangeli strains having the same geographical origin (Cuba Cuba et al. 1972, Cuba Cuba 1998, D'Alessandro 1976, D'Alessandro-Bacigalupo & Gore-Saravia 1992, 1999, Machado et al. 2001). Several T. rangeli strains have shown variable behaviour in different Rhodnius species; for example, strains from Colombia and Costa Rica do not produce metacyclic tripomastigotes in R. pallescens or R. ecuadoriensis. On the other hand, R. neglectus does not present salivary gland invasion when infected with strains from Panama or Costa Rica (D'Alessandro 1976). We have recently observed that T. rangeli strains isolated from R. colombiensis do not develop in the salivary glands of R. prolixus (Vallejo et al. 2002). PATHOGENICITY TO THE INSECT VECTOR The pathological effect of T. rangeli on R. prolixus was first observed by Grewal (1956, 1957). The author observed an increased amount of haemolymph and moulting deformities leading to high mortality. Other workers have confirmed different degrees of pathogenicity of T. rangeli for its vectors (Tobie 1964, 1965, Gomez 1967, Marinkelle 1968b, Watkins 1971a, b). MULTIPLICATION IN A VERTEBRATE HOST A question which continues to be the object of controversy is whether T. rangeli really does present some form of reproduction in a vertebrate host. In this sense, UrdanetaMorales and Tejero (1985) have described parasitaemia 7 times greater than that of the inoculum applied experimentally in lactating mice. A significant increase in the number of sanguineous tripomastigotes, following inoculation with a defined number of flagellates, suggests that T. rangeli probably multiplies within the vertebrate. Several researchers have sought in vain for forms of this parasite in the tissue of experimentally infected animals. For example, each tissue of mice infected with T. rangeli were histologically cross-sectioned; no tissue lesion or intracellular form was found in the 150 mice examined (HerbigSandreuter 1955). Grisard et al. (1999) recently studied several T. rangeli strains' interaction with mouse peritoneal macrophages, VERO, L929 and J77413 murine promonocyte cells, observing that less than 10% of such cells presented intracellular forms similar to those of amastigotes. These parasites disappeared 2 to 5 days after being infected without new free flagellates appearing in culture. Contrary to the forgoing, there are reports in the literature indicating the presence of reproductive T. rangeli forms in vertebrates (Grogl & Kuhn 1984, Scorza et al. 1986, Urdaneta Morales & Tejero 1986, Osorio et al. 1995). UrdanetaMorales and Tejero (1986) have described finding T. rangeli amastigote forms in cross-sections of heart, liver and spleen taken from NMRI lactating mice inoculated with forms of Venezuelan Perro82 strain culture. Osorio et al. (1995) demonstrated the presence of T. rangeli amastigotes in human U937 fibroblast culture. Such reports present the possibility that T. rangeli reproduction in a vertebrate host could depend on the characteristics of that strain being used. The reproductive phase of T. rangeli in the vertebrate host is currently not known and the mentioned observations need to be confirmed using cloned strains of T. rangeli to eliminate any possibility of contamination with T. cruzi. It has been observed that failure to experimentally infect susceptible mammals with T. rangeli has been related to the loss of flagellates' infectivity when they are kept in vitro for a long time. This phenomenon has been observed even when flagellates are formed in proven triatomid vectors' salivary glands, but (for as yet unknown reasons) the metatripomastigotes become non-infective (D'Alessandro 1976). According to the foregoing, it has thus become considered that T. rangeli strains must be studied as soon as possible after their isolation, before some of their main biological characteristics can become modified. Alternatively, several researchers have designed mechanisms for cyclically maintaining T. rangeli in a laboratory, by means of mousetriatomineculture, serial passages (Tobie 1964, 1968, D'Alessandro 1972, Cuba Cuba 1975). Vallejo et al. (1986) developed a maintenance system for maintaining T. rangeli strain infectivity, having serial passages each three months in culturetriatomine-mouseculture by means of which several strains have been kept for 20 years without losing their infectivity for vertebrate hosts. BIOCHEMICAL CHARACTERISTICS Several biochemical methods have been used for characterizing T. rangeli. The most promising have been: sensitivity to lysis mediated by the complement; characterizing surface sugars by using lectins; neuraminidase production; antigenic characterization; and isoenzyme analysis. By contrast with T. cruzi culture epimastigotes, T. rangeli culture epimastigotes are resistant to complement lysis activated by alternate pathway. This difference in susceptibility to lysis mediated by the complement has been used to differentiate the 2 species in culture or at vector intestine level (Schottelius & Muller 1984, Marinkelle et al. 1985). Several researchers have characterized T. cruzi and T. rangeli surface sugars (Schottelius & Muller 1984, Miranda Santos & Pereira 1984, Schottelius et al. 1986, Marinkelle et al. 1986, Schottelius 1989). The results of this body of research have shown several lectins' specific affinity for Dgalactose, Nacetylgalactosamine, N-acetylglucosamine and Nacetylneuraminic acid sugars on the surface of T. cruzi epimastigotes. The same researchers have found that T. cruzi and T. rangeli culture epimastigotes present mannose and glucose on their cell surface, presenting common agglutination with Canavalia ensiformis, Pisum sativum and Lens culinaris lectins. Miranda Santos and Pereira (1984) have reported specific Vicia villosa affinity (N-acetylgalactosamine affinity) for T. rangeli culture epimastigotes. Ongoing research indicates that T. cruzi has greater surface sugar abundance and diversity than that reported for T. rangeli. González et al. (1996) assayed extracts from 176 species of Colombian plant seeds and haemolymph from 16 native invertebrate species; they found that 7 seed extracts and 1 type of haemolymph agglutinated to T. cruzi and T. rangeli and 5 extracts agglutinated exclusively to T. cruzi, but none of the extracts agglutinated exclusively to T. rangeli. NEURAMINIDASE PRODUCTION Pereira and Moss (1985) reported that T. rangeli culture epimastigotes produced neuraminidase in easily detectable quantities (compared to T. cruzi culture epi-mastigotes). This enzyme thus becomes an important biochemical marker leading to the 2 species of flagellates being able to be differentiated. Vallejo and Marinkelle (1992) used 3 methods for detecting the presence of neuraminidase in the promesenteron, postmesenteron, rectal ampulla, haemolymph and salivary glands of 600 R. prolixus experimentally infected with T. rangeli San Agustín. Haemoglutination methods using peanut lectin (Arachis hypogaea), fluorescence reaction with peanut lectin conjugated with fluorescein isothiocyanate and fluorescence emitted by 4methyl-umbeliferone showed neuraminidase presence in all cases in T. rangeli culture supernatant in Tobie medium having 8 to 15 days growth. None of the 3 methods detected neuraminidase presence in R. prolixus infected with T. rangeli, indicating that this enzyme is probably not produced in vivo or was produced in non-detectable quantities by the methods used. Different studies have shown that T. rangeli presents a crossed antigenic reaction with T. cruzi. This crossed reaction has been shown by immunoelectrophoresis, ELISA, indirect immunofluorescence, immunoblotting, double immunodiffusion (Afchain et al. 1979, Antony et al. 1979, Guhl & Marinkelle 1982, Grogl & Kuhn 1984, Basso et al. 1989, O' Daly et al. 1994, Saldaña & Sousa 1996). These studies have generally shown that the 2 parasite species share at least 60% of their antigens. Other studies have also pointed to an apparent diminution of the severity of Chagas disease symptomatology when there is mixed infection by the 2 parasite species (Guhl et al. 1985, Hudson et al. 1988). ANALYSIS OF ISOENZYMES IN T. RANGELI Analysis of several hundred T. cruzi strains isolated in different Latin-American countries has led to information being obtained regarding this parasite's genetic structure. On the contrary, few isoenzyme studies have been done on T. rangeli; also, only a small number of strains have been used in each study. A first approach using enzymes for studying T. rangeli was reported by Kreutzer and Sousa (1981); they described differences in isoenzyme profiles for strains from Panama. Ebert (1986) studied T. rangeli isoenzymes and their relationship to other trypanosomes transmitted by triatomids. This author found that 3 out of the 9 enzymes examined showed T. rangeli-specific profiles (NSE, PGM and GP1). He also found little enzymatic variation for the 14 strains examined. Holguin et al. (1987) later analyzed T. rangeli strains from wild and domestic cycles in Colombia. When the above authors were studying 14 enzymes (GP1, PEP, MPI, PGM, ALAT, ASAT, ACON, MDH, KP: kinase piruvate, ICD, G6PD, LDH, KGP: kinase phosphoglycerate, ME) they found that only the ICD enzyme showed some variation, indicating that the isoenzymes did not manage to differentiate domestic strains from the wild ones, contrary to that found for T. cruzi. Acosta et al. (1991) differentiated T. cruzi from T. rangeli from Honduras by using 4 methodologies (lysis susceptibility by complement, lectin reactivity, reactivity to monoclonal T. cruzi-specific antibodies and isoenzyme electrophoretic patterns). An important contribution was made towards the isoenzymatic characterization of T. rangeli by Steindel et al. (1994) who used 16 T. rangeli strains; 8 of these were isolated from R. prolixus and patients from Honduras, Colombia and Venezuela, whilst another 8 came from P. megistus or rodents (Echimys dasythrix) from the state of Santa Catarina in Southern Brazil. These authors studied 5 enzymes (ALAT, ASAT, GP1, PGM, ME), finding that all the Santa Catarina strains were isoenzymatically identical. Those strains from Honduras, Colombia and Venezuela also formed a highly homogeneous group. The 2 groups showed different patterns for all enzymes, except for the malic enzyme. This suggested the existence of 2 different groups of T. rangeli strains. Triana et al. (1999) recently studied the genetic variability of T. cruzi and T. rangeli strains isolated from different hosts. They found low variability in 3 T. rangeli strains when using electrophoresis on 15 systems' enzymatic and analyzed DNA (kDNA) digested with endonucleases. MOLECULAR KARYOTYPES Analysis of several T. rangeli strains' molecular karyotypes has shown variation in the size of the chromosomic bands ranging from 400 Kb to 2,900 Kb, revealing chromosomic polymorphism between strains isolated from El Salvador, Honduras, Venezuela, Colombia and Panama and strains from Southern Brazil (Henriksson et al. 1996, Toaldo et al. 2001). KINETOPLAST DNA (kDNA) During the last 20 years, kDNA sequences have been used for tripanosomatid diagnosis and characterization. This extranuclear DNA forms a compact network within a mitochondrial complex. kDNA represents these parasites' mitochondrial genome, constituting between 10% and 20% of cellular DNA. The kDNA network has between 5,000 and 10,000 minicircles whose size varies from 0.5 Kb (Trypanosoma vivax) to 2.5 Kb (Crithidia fasciculata) and has 25 to 50 maxicircle copies varying between 19 Kb (Bodo caudatus) and 39 Kb (Phytomonas davidii) (Vallejo 1998). The maxicircles are similar to higher eukaryote mitochondrial DNA, codifying ribosomal RNAs and proteins involved in translating energy in the mitochondria (Simpson 1987). The presence of large quantities of minicircles reflects these organisms' ability to revise mitochondrial transcripts' nucleotide sequences by a new process called RNA editing, in which several uracil residues are inserted or eliminated at specific sites in the sequence (Stuart & Feagin 1992). Linearized minicircles can be easily observed when tripanosomatid total DNA or kDNA is digested with some restriction enzymes. For example, regarding T. rangeli, total DNA digestion with Hae III allows bands corresponding to 1.8 and 1.6 Kb to be visualized in agarose gels (Vallejo et al. 1994). All minicircles present at least one conserved or minirepeat region varying between 100 and 200 bp. Each tripanosomatid specie has the same number of conserved regions. Only one conserved region has been observed in C. oncopelti, T. brucei and Leishmania tarentolae. There are 2 copies of the conserved region in C. fasciculata, T. lewisi and Phytomonas serpens organized as direct repetitions, located at 1,800 bp from each other. There are 4 copies of the conserved region in T. cruzi organized as direct repetitions, located at 90o. It is worth stating that minicircles have been observed in the same T. rangeli strain or clone having 1, 2 and 4 conserved regions (Recinos et al. 1994, Vallejo et al. 1994). Comparing T. cruzi and T. rangeli minicircle conserved regions reveals high homology in conserved sequence block (CSB1, CSB2 and CSB3) sequence, order and distance. Primers have been designed on these blocks for detecting T. cruzi or T. rangeli by using PCR. The S35 and S36 primers have amplified a 330 bp fragment with high sensitivity leading to 0.015 fg or 10 parasite minicircles (Sturm et al. 1989) or a parasite in 20 ml of blood being detected. These same primers were used recently to detect T. cruzi in mummified tissue from Northern Chile (Guhl et al. 1997). These primers also react with T. rangeli minicircles, allowing amplification of a 760 bp fragment derived from 1,600 bp minicircles having two conserved regions and a heterogeneous set of fragments from 300 to 450 bp derived from 1,600 bp minicircles from 4 conserved regions (Vallejo et al. 1999). Vallejo et al. (2002, 2003), designed a duplex polymerase chain reaction (PCR) reaction using 3 primers (S35/S36/KP1L) allowing amplification from minicircles with a conserved region (denoted as KP1), 2 conserved regions (denoted as KP2) and 4 conserved regions (denoted as KP3). Strains isolated from R. prolixus presented K1, KP2 and KP3 minicircle amplification products. On the other hand, strains isolated from R. colombiensis R. pallescens or P. megistus presented amplification products derived from KP2 and KP3 minicircles, but not from KP1. Vallejo et al. (2002), examined 150 R. colombiensis adults, finding T. rangeli KP1(+) and KP1(-) in the intestine of several specimens and T. rangeli KP1(-) in salivary glands in 15 individuals, indicating that in natural infections, KP1(+) strains can be found in the intestine but these strains do not invade R. colombiensis haemolymph and salivary glands. The authors observed a similar situation in 90 examined R. prolixus where it was possible to find T. rangeli KP1(-) and KP1(+) in the intestine of several specimens, T. rangeli KP1(+) only being detected in the salivary glands of 12 individuals. The foregoing data suggests that some Rhodnius genus species are susceptible to T. rangeli KP1(+) strains and others to KP1(-), so that each Rhodnius species would select the sub-population to which it is susceptible in nature to transmit it by salivary inoculation to the vertebrate host. NUCLEAR DNA Macedo et al. (1993) carried out a minisatellite study (VNTR) using a 33.15 probe. The multiband pattern which they obtained allowed T. cruzi to be differentiated from T. rangeli. The dendrogram obtained revealed the existence of two groups, one formed by T. rangeli strains from Central America, Colombia and Venezuela and another formed by T. rangeli strains from Southeastern Brazil (Santa Catarina). RANDON AMPLIFIED POLYMORPHIC DNA (RAPD) Few studies have been done using APPCR for T. rangeli characterization. In a first study, analysis of 138 RAPD bands generated by 6 primers showed that 2 T. rangeli strains from Santa Catarina formed a group separated from another group of 8 T. rangeli strains isolated from Honduras, Colombia and Venezuela (Steindel et al. 1994). HISTONE GENES AND OTHER REPETITIVE ELEMENTS The gene encoding H2A histone was isolated and characterized from the T. rangeli C23 strain (Puerta et al. 2000). It was observed that this gene was organized into tandem repetitions, located in a 1.9 Mb chromosome; it transcribed 0.7 kb mRNA. At least 10 different nuclear DNA repetitive elements have been described in T. cruzi (Requena et al. 1996). On the contrary, few T. rangeli nuclear DNA repetitive elements have been reported in the literature. P542 is an element possessing 103 copies per haploid genome and is found to be distributed throughout several chromosomic bands. The element has been sequenced in an attempt to design primers for detecting the parasite's presence. Preliminary studies have shown that this element is very useful for detecting T. rangeli in the intestine of vectors naturally infected with mixed T. cruzi and T. rangeli infections (Vargas et al. 2000). GENETIC CHARACTERIZATION OF T. RANGELI Little is known about the genetic structure of T. rangeli; few studies have analyzed multiple loci (isoenzymes and RAPDs), thus limiting genetic studies regarding this parasite's populations. Vallejo et al. (2003), recently observed a non-random association between mitochondrial kDNA amplification and nuclear mini-exon gene, indicating dimorphism within both DNA sequences: KP1, KP2 and KP3 or KP2 and KP3 products for kDNA and 380 bp or 340 bp products for the mini-exon. Either KP1, KP2 and KP3 kDNA product association with the 340 bp mini-exon product or KP2 and KP3 kDNA product association with the 380 bp mini-exon product was observed within individual strains. On the other hand, several T. rangeli strains from distant geographical origins have been shown to have identical kDNA (KP2 and KP3) and mini-exon profiles (380 bp), indicating that kDNA and mini-exon genotypes are replicated as a unit, without recombination and that this strict correlation between the markers persists over distant localities. Based on these results, the authors propose two alternative hypotheses, considering that T. rangeli could have clonal evolution or have been subjected to speciation processes. However, further work must be done with populations having different geographical origins, a greater number of strains must be used and population genetic analysis within each of the two groups, using a multi-locus approach, could lead to resolving the difference between the two hypotheses. Statistical tests of population genetic data should be done and if there is still a linkage disequilibrium after such analysis, then this favours clonal evolution rather than cryptic speciation. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03097t1.jpg] [oc03097t2.jpg] |
| |||||||||

{kind=link}
{kind=link}