 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 4, June, 2003, pp. 487-493 Characterization of [Ca2+]i Responses in Primary Cultures of Mouse Cardiomyocytes Induced by Trypanosoma cruzi Trypomastigotes Luciana R Garzoni, Masako Oya Masuda*, Márcia M Capella**, Anibal Gil Lopes**, Maria de Nazareth S Leal de Meirelles/+ Laboratório de Ultra-estrutura Celular, Departamento de Ultra-estrutura e Biologia Celular, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil *Laboratório de Eletrofisiologia Cardíaca **Laboratório de Fisiologia Renal, Instituto de Biofísica Carlos Chagas Filho, UFRJ, Rio de Janeiro, RJ, Brasil This research received financial
support from Conselho Nacional de Desenvolvimento Científico e Tecnológico
(CNPq), Programa de Apoio à Pesquisa Estratégica em Saúde
(Papes-Fiocruz), Fundação de Amparo à Pesquisa do Estado
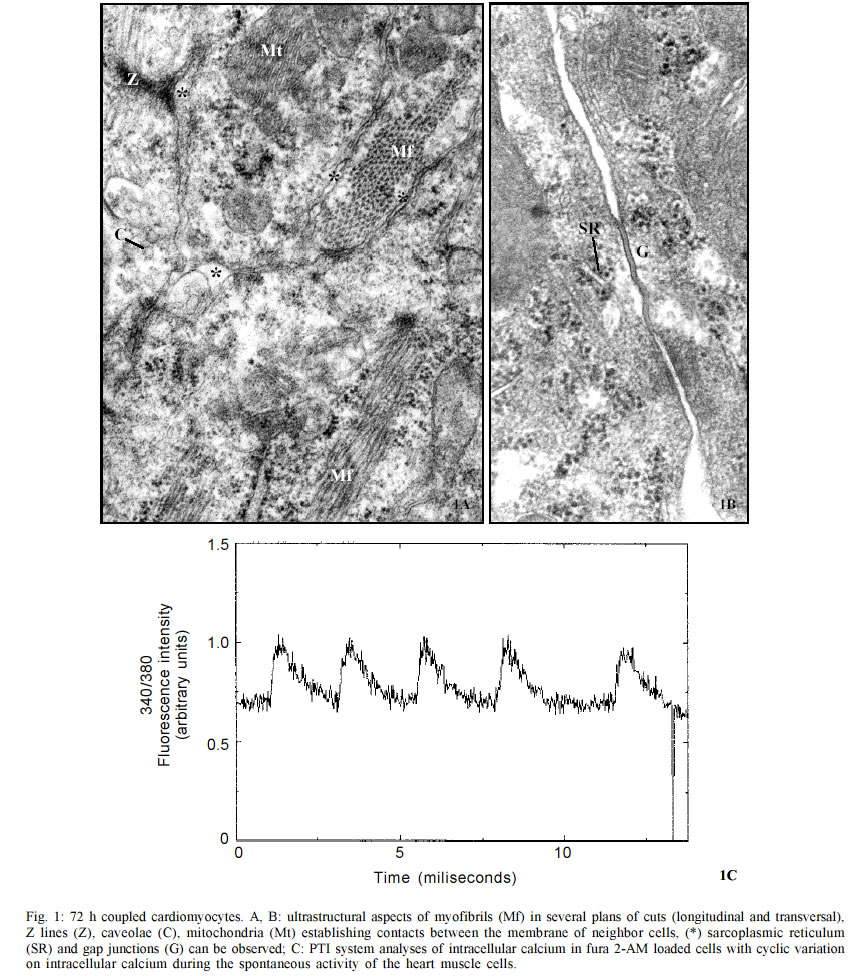
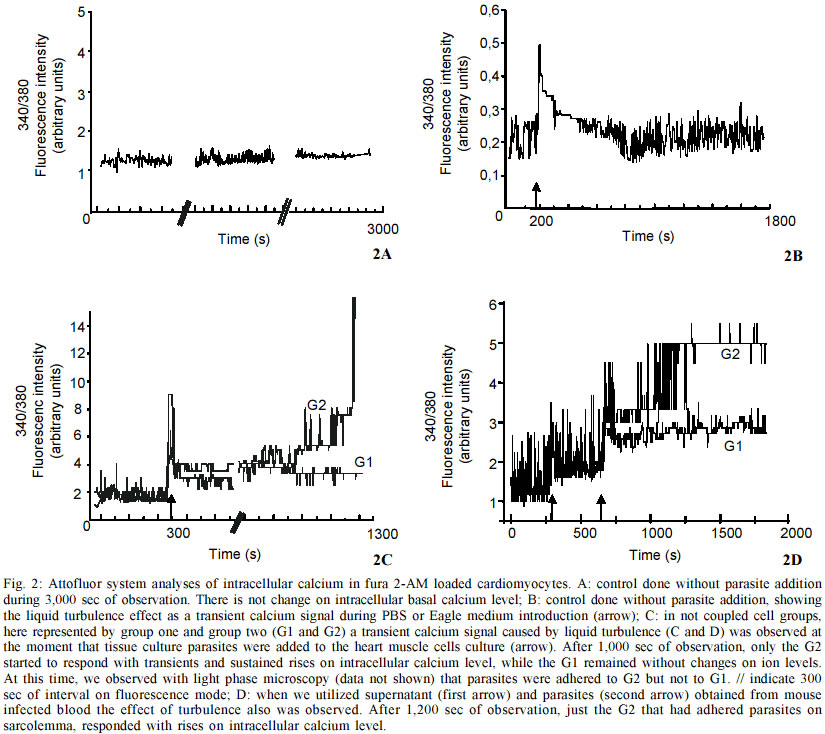
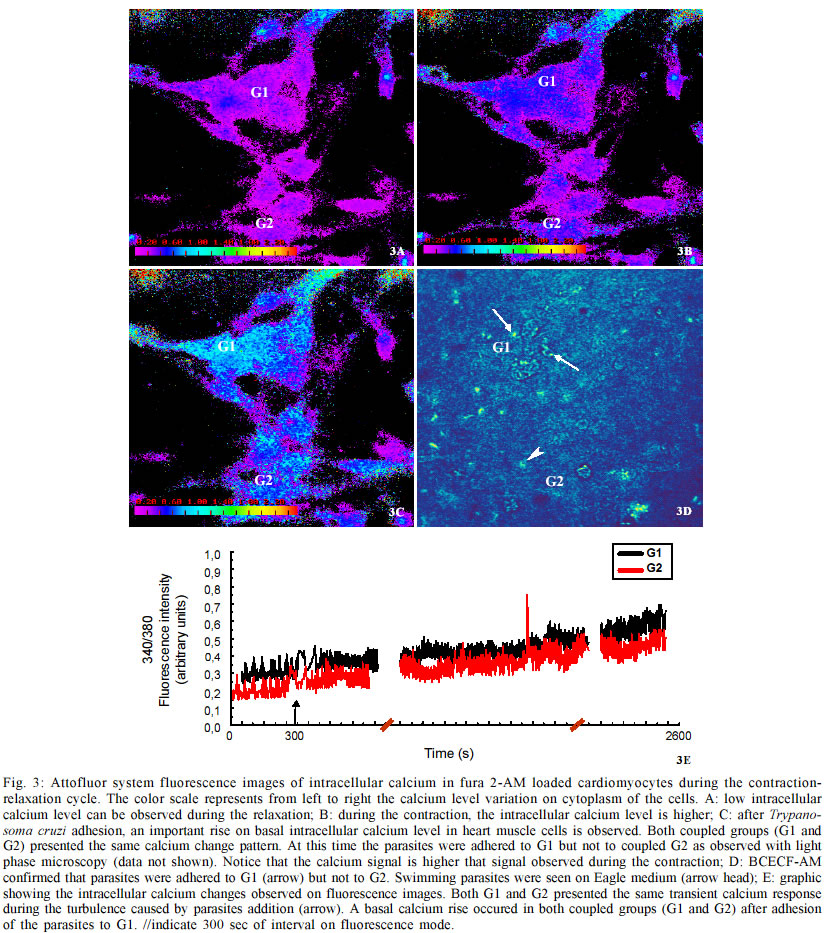
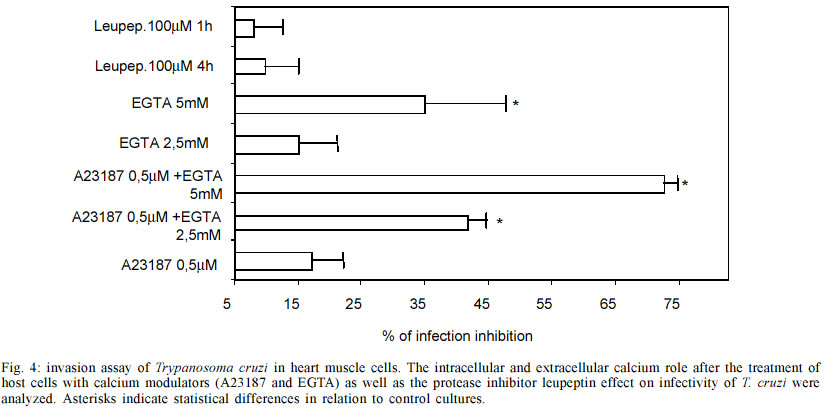
do Rio de Janeiro (Faperj). Received 15 October 2002 Code Number: oc03106 Trypanosoma cruzi, the protozoan responsible for Chagas disease, employs distinct strategies to invade mammalian host cells. In the present work we investigated the participation of calcium ions on the invasion process using primary cultures of embryonic mice cardiomyocytes which exhibit spontaneous contraction in vitro. Using Fura 2-AM we found that T. cruzi was able to induce a sustained increase in basal intracellular Ca2+ level in heart muscle cells (HMC), the response being associated or not with Ca2+ transient peaks. Assays performed with both Y and CL strains indicated that the changes in intracellular Ca2+ started after parasites contacted with the cardiomyocytes and the evoked response was higher than the Ca2+ signal associated to the spontaneous contractions. The possible role of the extracellular and intracellular Ca2+ levels on T. cruzi invasion process was evaluated using the extracellular Ca2+ chelator EGTA alone or in association with the calcium ionophore A23187. Significant dose dependent inhibition of the invasion levels were found when intracellular calcium release was prevented by the association of EGTA +A23187 in calcium free medium. Dose response experiments indicated that EGTA 2.5 mM to 5 mM decreased the invasion level by 15.2 to 35.1% while A23187 (0.5 µM) alone did not induce significant effects (17%); treatment of the cultures with the protease inhibitor leupeptin did not affect the endocytic index, thus arguing against the involvement of leupeptin sensitive proteases in the invasion of HMC. Key words: Trypanosoma cruzi - cardiomyocytes - transients and sustained [Ca2+]i rises Trypanosoma cruzi is the protozoan parasite responsible for Chagas disease (Chagas 1909), an important public health problem in Latin America. A total of 16-18 millions people are infected by this parasite in the world (WHO 1997) and the cardiomyopathy is a serious consequence of chronic infection. The parasite is transmitted to man and to other vertebrate hosts by the triatominae insect of the Reduviidae family. Metacyclic trypomastigote forms invade macrophages, cardiac and other vertebrate cells and differentiate to round and multiplicative amastigote forms. After successive intracellular divisions, amastigotes transform to trypomastigotes. After host cell burst, the flagelates are released and propagate the infection via the bloodstream. Mouse models are reliable tools for studies of Chagas heart disease (Melo & Brener 1971, Grimaud & Andrade 1984). Primary culture of heart muscle cells (HMC) are an useful model to study T. cruzi-cardiomyocyte interaction (Meirelles et al. 1987, 1992, Pereira et al. 1993, Soeiro et al. 1995, Barbosa & Meirelles 1995). Following the normal myogenic process these cells attach to the gelatin-coated substrate and exhibit a typical bipolar aspect. After an initial period of proliferation, coupling between the cells is accomplished, as indicated by the synchronized contraction. Ultrastructural studies and electrophysiological approaches showed that these cardiac cells present the same basic structure found in mammalian heart cells in vivo such as specialized structures including sarcotubular system, intercalated discs, myofibriles, coupling interaction among the cells and spontaneous contractility (Meirelles et al. 1986, Aprigliano et al. 1993, Barbosa & Meirelles 1995). In coupled cardiomyocytes, endocytosis seems to be the main process leading to parasite invasion (Meirelles et al. 1986, Barbosa & Meirelles 1995). Early studies with irreversible synthetic inhibitors of cruzipain suggested that protease activity was required for HMC invasion (Meirelles et al. 1992). It has been described that T. cruzi induces cytosolic free calcium transients during the invasion process in myoblasts and fibroblast lineages, cells that do not present spontaneous activity (Moreno et al. 1994, Rodriguez et al. 1995). Ca2+ signaling activity was detected in soluble extracts of metacyclic trypo-mastigotes in response to the binding of the surface parasite molecules, i.e. gp82 or gp35/50, to HeLa cells receptors. These studies showed that metacyclic parasites trigger cytosolic Ca2+ transients without need of proteolysis involvement (Dorta et al. 1995). In other cell systems, it was shown that generation of cytosolic Ca2+ was induced by oligopeptidase B (Caler et al. 1998). Analysis of the requirement for Ca2+ signal induced by soluble try-pomastigote extracts indicated that it was blocked by leupeptin, a protease inhibitor or by specific antibodies to recombinant Oligopeptidase B (Caler et al. 1998). In primary culture of umbilical vein endothelial cells (HUVECs) or in Chinese hamster ovary cells overexpress-ing the B2 type of bradikinin receptor (CHO-B2R), the parasites relied upon cruzipain to elicit [Ca2+]i transients through B2 receptors (Scharfstein et al. 2000). Combined, these studies indicate that T. cruzi elicit calcium rises on host cell through various molecular pathways. In the present paper, we show that both intracellular and extracellular calcium are necessary for parasite invasion, and that the parasites were able to induce both transient and sustained rises on the [Ca2+]i levels in HMC following adhesive contacts with the sarcolemma. MATERIALS AND METHODS Reagents - Fura 2-AM [Fura-2 (acetoxymethyl) ester] and BCECF-AM [2',7'-Bis(2-carboxyethyl)-5(6)-carboxyfluorescein tetra (acetoxymethyl) ester] were obtained from Molecular Probes, Inc. (Eugene, Oregon, USA) and EGTA [Ethylene glycol-bis(2-aminoethyl)-N,N,N',N'-tetraacetic acid], calcium Ionophore A23187, DMSO (Dimethyl sulfoxide) and Leupeptin, were purchased from Sigma Chemical Co. (St. Louis, MO, USA) Host cell and parasites - Hearts of 18 days old mouse embryos of Swiss mice were submitted to mechanical and to enzymatic dissociation using trypsin 0.05% and 0.01% collagenase in phosphate buffer saline (PBS) at 37°C, following the method previously described (Meirelles et al. 1984, 1986). The ventricular heart muscle cells were plated in 0.02% gelatin-coated plastic bottle for obtention of cultures, in glass coverslip maintained in 24 well plates to be used for invasion assays, or in a special chamber of plastic Petri dish containing a 1 cm central well covered by a thin glass coverslip (0.7 mm) mounted with silicone glue for the digital imaging fluorescence microscopy. The cells were maintained at 37°C in 5% of CO2 atmosphere in Dulbecco' modified Eagle medium (DMEM) (Dulbecco & Freeman 1959), supplemented with 5% fetal calf serum, 1 mM CaCl2, 15% horse serum, 1 mM L-glutamine, 2% chick embryo extract, 1000 U/ml penicillin and streptomycin 50 µg/ml-1. For ultrastructural studies, HMC were fixed in glutaraldehyde and osmium tetroxide and embbeded in Epon. Trypomastigote forms of T. cruzi Y and CL stocks were used throughout the experiments. The parasites were obtained from the supernatant of HMC previously infected for 24 h at 37°C employing a parasite:cell ratio of 10:1 at 37°C. After 72-96 h of infection trypomastigote forms liberated in culture medium were collected for the invasion experiments. Parasites obtained from blood of infected Swiss mice were also used. The animals were inoculated with 104 trypomastigote forms and after seven days, at the peak of parasitaemia, heart puncture was done and the blood was processed for obtention of the parasites. Trypomastigote forms were then maintained in DMEM without serum, at 4°C overnight. Before each experiment, the culture or blood parasites were submitted to centrifugation at 2,000 g, the supernatant was colleted, the parasites were washed three times in PBS, ressuspended in DME and added to HMC at a ratio of 20:1 (parasite: cell) in a final volume of 300 µl. Measurement of cytosolic calcium in HMC - 15 X 104 HMC were platted for 72 h into a well localized in the special chamber with coverslip bottom as described above. After this time, the cells were already completely differentiated and we found groups of 4-6 or more coupled cardiomyocytes displaying regular spontaneous contraction (Meirelles et al. 1986, Aprigliano et al. 1993). HMC were then washed in DMEM, and loaded for 30 min at 37°C with Fura 2-AM 4 µg/ml. Then the cells were washed to remove the free extracellular dye and were maintained in DMEM supplemented with 1mM CaCl2 during the whole experiment. Changes on basal level of intracellular calcium analysis were performed in an Axiovert 100 microscope equipped with a system of exchange excitation filter (334 and 380 nm) as well as a digital ratio imaging system (Attofluor, Oberkochen,Germany, Zeiss), under 63X oil objective. Fluorescence images were collected by a digital CCD camera using 510 nm filter. [Ca2+]i were monitored during 2,000 sec at 37°C in both single and clusters of HMC. Fluorescence images were digitalized and a visual display of 334/380 nm ratio at 0.5 points/sec sampling rate was produced. Attograph software was used to generate the graphics representing [Ca2+]i variations. The background, non stimulated changes in [Ca2+]i was monitored for 3,000 sec. The effect of the turbulence upon addition of the DMEM or PBS on the fluorescence signal lasted for few seconds. The interaction of HMC with the parasites were checked under phase contrast optical microscopy in order to improve the parasites visualization; in some experiments they were previously loaded with 4 µg/ml BCECF, which excited at 490 nm and emitted at 510 nm. The spontaneous activity of cardiomyocytes was performed using a Nikon fluorescence microscopy and the PTI system (Photon Technology Inc). Fluorescence signal of 334/380 nm ratio was obtained at 100 points/sec sampling rate. The Attograph software was used to generate tracings representing [Ca2+]i transients of individual or groups of cells, as well as average responses of 5 to 8 cell groups (20-30 cells) in each experiment (n = 11). Invasion assay - HMC were plated in the 24 well plates containing round coverslips at a density of 15 X 104 cells/well. After 48 h of cell culture, the contracting cardio-myocytes were pre-treated with DMEM containing the extracellular calcium chelator EGTA, the ionophore A23187 or both calcium modulators EGTA + A23187. The HMC pretreatments were done at 37°C, as follows: 10 min with 2.5 mM or 5 mM EGTA, 5 min with 0.5 µM A23187, 5 min with 2.5 mM EGTA plus 0.5 µM A23187, or 5 mM EGTA plus 0.5 µM A231870. In all cases, HMC were washed three times with PBS and then the trypo-mastigotes forms of T. cruzi (Y strain) were added to HMC monolayer at parasite:cell ratio multiplicity of 10:1, at a final volume of 300 µl. After 3 h of interaction at 37°C, the free parasites were removed by three washes with PBS and the infected cardiomyocyte cultures were fixed with Bouin and stained in Giemsa solution. We tested the effect of the protease inhibitor leupeptin on invasion of HMC by T. cruzi, treating for 1 h and 4 h both parasites and HMC with 10 and 100 µM with this inhibitor in DMEM. Controls were performed by incubating HMC cultures with DMEM containing the same final concentration of DMSO, which was the A23187 and leupeptin solvent used in these assays. Invasion was measured by counting the number of infected HMC in a total of 200 cells per coverslip. Triplicate for each variable was done and the percentage of invasion was calculated.Values represent means ± standard deviation of three independent experiments. Statistical analysis was done using the Student t test and p < 0.05 values were considered significant. RESULTS Ultrastructural analyses of 72 h coupled HMC cells revealed the presence of caveolae (C), Z lines (Z), mitochondria (Mt) and myofibrils (Mf) in longitudinal and transversal plans of cut (Fig. 1A). In addition, sarcoplasmic reticulum (SR), gap junctions (G) were also observed by electron microscopy (Fig. 1B). These cells displayed spontaneous contractile activity activity, with the intracellular calcium levels changing cyclically as observed in Fura-2 AM loaded cells at PTI system (Fig. 1C). Changes on basal intracellular calcium level were analyzed with Attofluor system for 2,200 sec. Non infected cells showed no significant alteration in the mean basal [Ca2+]i level (Fig. 2A). Controls with PBS or culture medium without parasites induced transient and basal [Ca2+]i increase immediately, which stabilized after a few seconds (Fig. 2B). The same pattern of response was observed upon addition of parasites originating from tissue culture (Figs 2C, 3E) or blood forms (Fig. 2D) or after parasite supernatant addition, suggesting that it resulted from turbulence effects. We then followed the effect of T. cruzi invasion on [Ca2+]i responses in individual cells. We observed that two non coupled groups of HMC, G1 and G2 displayed the turbulence effect upon the parasites addition (Figs 2C, 2D-arrow). In one case, G2, we observed a transient [Ca2+]i response associated to a sustained increase of basal [Ca2+]i , starting at 1,700 sec. At this moment, phase light microscopy (not shown) indicated that the parasites were only adhered to G2 group. In the G1 group of cells, the fluorescence signal stayed stable and under these conditions, no parasite were observed adhered or even in proximity of cells (Fig. 2C). Fluorescence microscopy examination revealed changes on the intracellular calcium levels during the contraction-relaxation cycle in coupled cardiomyocytes, as well as during the invasion of T. cruzi. Differences on intracellular calcium levels were detected during the relaxation (Fig. 3A) and contraction (Fig. 3B) cycle of HMC. After the parasite addition, some HMC groups started to show rises on intracellular calcium (G1 and G2 _ Figs 3C, 3E). With phase microscopy we could observe parasites adhered to responsive G1 (not shown). This was confirmed with fluorescence images of the BCECF-AM loaded parasites (Fig. 3D). The group G2 (Fig. 3C) that did not show adhered parasites, but was coupled to parasite responsive group G1, presented the same pattern of change on intracellular calcium level observed in G1. Assays performed with bloodstream forms of the Y and CL (not shown) strains (Fig. 2D) or with tissue culture (Figs 2C, 3C) parasites induced the same rises of intracellular calcium response, following adhesion to HMC sarcolemma. In all experiments the changes in intracellular Ca2+ in HMC induced by T. cruzi were higher than the contraction Ca2+ signal (Figs 3B, 3C). Alterations on infection levels of T. cruzi in HMC were investigated after treatments with the protease inhibitor leupeptin or with calcium modulators. Addition of leupeptin during the interaction of parasites and HMC did not show any significant effects over control cultures (Fig. 4). In contrast, treatment with calcium modulators showed alterations in both intracellular and extracellular levels of Ca2+. Moreover, these compouds affected the outcome of infection by T. cruzi. The depletion of host cell intracellular calcium before the interaction with parasites by treatment the cultures with Ca2+ ionophore A23187 plus EGTA inhibited the parasite entry. When intracellular calcium was depleted with 2.5 mM or 5 mM EGTA plus 0.5 µM ionophore A23187, a significant dose dependent inhibition (41.7% and 72%) were observed, respectively. Treating the cells exclusively with EGTA 2.5 mM had no significant effect on infection (15%) while treatment with 5 mM of the chelator inhibited 35% of invasion. Addition of A23187 alone induced no significant inhibition level of 17%. Pre-treatment with DMSO had no effect on the T. cruzi invasion level in HMC. DISCUSSION In our HMC culture system the cells are coupled (Aprigliano et al. 1993) and present spontaneous contractile activity. The coupling between the cells assures the fast transmission of electric impulse, resulting in synchronic contraction of the cardiomyocytes. In the present work, we showed cyclic variations on the intracellular calcium levels during the contraction and relaxation, while exposure to T. cruzi trypomastigotes induce transient rises followed by sustained increase in the basal levels of [Ca2+]i. At the moment when the parasites were introduced into the chamber, liquid turbulence induced increases in [Ca2+]i in cardiomyocytes, suggesting that extracellular Ca2+ influx occurred through membrane Ca2+ channels that are sensitive to mechanical stress (Pickard & Ding 1993). Nevertheless, [Ca2+]i rises were repeatedly observed in cardiomyocytes as soon as the parasites adhered to the sarcolemma. The increase in the intracellular calcium was shared by the coupled cells, probably through gap junctions. Trypomastigotes loaded with BCECF-AM could be easily visualized attached to cardiomyocyte surface. Calcium responses are critically required for invasion of cardiomyocytes as shown with other cell types (Osuna et al. 1990, Moreno et al. 1994, Tardieux et al. 1994, Dorta et al. 1995, Meirelles et al. 1999, Scharfstein et al. 2000). Thapsigargin sensible sarcoplasmic reticulum Ca2+- ATPase (SERCA) was involved in T. cruzi invasion on coupled cardiomyocytes since the treatment of HMC with this drug inhibited parasite invasion (Meirelles et al. 1999, Todorov et al. 2003). Treatment with calcium modulators showed the importance of both host cell intracellular calcium as well as of extracellular calcium during the invasion of cardiomyocytes by T. cruzi. Meirelles et al. (1992) have shown that Z-(SBZ)Cys-Phe-CHN2, a membrane permeable diazomethylketone inhibitor of cruzipain, partially blocked T. cruzi invasion of cardiomyocytes. Barr et al. (1996) reported that a leupeptin-sensitive Oligopeptidase B (Tardieux et al. 1994, Burleigh & Andrews 1998, Caler et al. 1998) present in a soluble fraction of trypomastigotes produced fast repetitive cytosolic Ca2+ transients, each one associated with cell contraction. The data reported in the present work does not support a role for leupeptin-sensitive peptidases in this process. Dorta et al. (1995) have reported another case of Ca2+ signaling activity independent of Oligopeptidase B activity while studying soluble extracts of T. cruzi metacyclic trypomastigote during HeLa cells invasion. Recent studies by Scharfstein et al. (2000) offered an alternative explanation for the negative data obtained with leupeptin. Using host cells that express G-protein coupled kinin-receptors, they have shown that parasite invasion was enhanced due to the parasite' ability to generate kinins through the proteolytic activity of cruzipain. Once released, the kinin agonist elicits vigorous [Ca2+]i transient due to activation of B2- kinin receptor (Scharfstein et al. 2000). In the same study, they showed that membrane-permeable cruzipain inhibitors, such as Z-(SBZ)Cys-Phe-CHN2, but not hydrophylic inhbitors, blocked the invasion process. The authors concluded that cruzipain _ which like Oligopeptidase B is sensitive to leupeptin _ must release the kinin agonist in a secluded spaces formed by the juxtaposition of host and parasite plasma membranes. Thus, it is possible that leupeptin, a water soluble inhibitor, is unable to reach sites where cruzipain or oligopeptidase generate the [Ca2+]i signaling mediators required for parasite invasion. In summary, our data suggests that activation responses evoked by T. cruzi trypomastigotes disturb calcium homeostasis in HMC. Parasite persistence in the heart from chronically infected patients may provoke repetive activation of myocardial cells, perhaps in the long term contributing to the pathogenesis of Chagas disease. ACKNOWLEDGEMENTS To Bruno Ávila for the image technical assistance, to Alanderson da Rocha Nogueira for the technical support with the cultures and Núbia Gabriela de CB Chedid for the PTI analyses. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03106f3.jpg] [oc03106f2.jpg] [oc03106f4.jpg] [oc03106f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}