 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
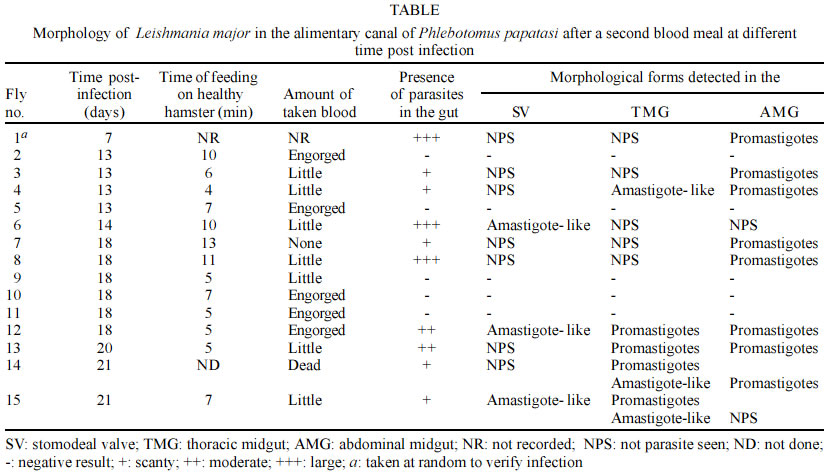
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 4, June, 2003, pp. 495-498 Detection of Amastigote-like Forms in the Valve of Phlebotomus papatasi Infected with Leishmania major Nestor Añez/+, Yinshan Tang*, Agustina Rojas, Gladys Crisante, Mireille Killick-Kendrick**, Robert Killick-Kendrick** Departamento de Biología,
Facultad de Ciencias, Universidad de Los Andes, Mérida, 5101, Venezuela
*14 Newquay Drive, Bramhall, Stockport, Cheshire, UK **2 Place du Temple,
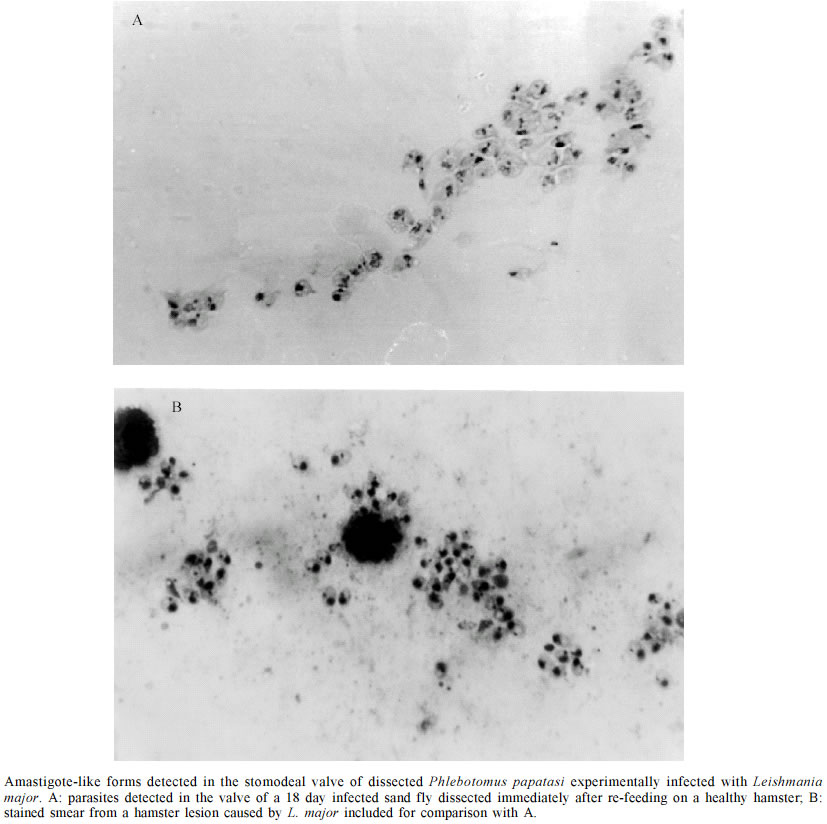
Sumène, France Financial support given by Fonacit Grant no. G-99000036, and CDCHT-ULA-C-1016-00-07-AA. Received 5 December 2002 Code Number: oc03107 A massive and homogeneous amount of amastigote-like forms was detected in the stomodeal valve (SV) and the thoracic mid-gut (TMG) of Leishmania major-infected Phlebotomus papatasi, which received a second blood meal 13 to 21 days post-infection on healthy anaesthetized hamsters. After re-feeding, the infected sand flies were dissected out to examine the morphology of the parasite in SV, TMG and the abdominal mid-gut (AMG). Different promastigote forms were seen in the infected flies. Among these included typical promastigotes (nectomonads and haptomonads), paramastigotes, metacyclic promastigotes and, in some samples, the here-reported amastigote-like forms. The Leishmania amastigote-like forms were detected in the SV of sand flies with 14, 18 and 21 days of infection as well as in the TMG at 13 and 18 days post-infection. However, the amastigote-like forms were not detected in the AMG. Factors such as the acidic pH predominating the TMG and the SV, as well as the temperature of the ingested blood, among others, are suggested as contributing to the transformation of the typical promastigotes into the amastigote-like forms. The significance of this finding is discussed and the possible biological advantage for transmission of Leishmania is considered. Key words: Phlebotomus papatasi - Leishmania major - amastigote-like forms - transmission Parasites of the genus Leishmania exhibit, during their life cycle, two well-known forms. The flagellated and mobile promastigote form, which multiplies in the gut of phlebotomine sand fly vectors, and the intracellular non-motile amastigote form normally found within macrophage phagolysosomes of the vertebrate host (Killick-Kendrick 1990). The sand fly gut is an environment specialized for blood digestion as well as the location where Leishmania-parasites replicate and develop into the infective metacyclic forms that invade the vertebrate host (Tang et al. 1998). Thus far, the Leishmania metacyclic form has been described as a parasite possessing a short body, and a very long flagellum which is found in the foregut including the proboscis of infected sand flies. The metacyclic form is also complement resistant and able to transform into the amastigote form upon infecting the vertebrate host (Killick-Kendrick 1990). Recent studies have demonstrated that the combined effect of low external pH and temperatures are factors that induce in vitro metacyclogenesis of Leishmania (Bates & Tetley 1993, Zilberstein & Shapira 1994, Valencia-Pacheco et al. 1998). In addition, Tang et al. (1998) demonstrated that sand flies have an acidic pH in the thoracic mid-gut (TMG), which is about 6.8 or below when the crop is full of sugar or when a blood meal has been digested. These findings may help interpretation concerning morphological changes in the Leishmania-parasite population in experimentally infected sand flies. In the present paper we report the detection of amastigote-like forms in the stomodeal valve (SV) and TMG of colonized Phlebotomus papatasi which received a second blood meal 13 to 21 days after an experimental infection with Leishmania major. MATERIALS AND METHODS Sixty eight specimens of Ph. papatasi from a laboratory colony were experimentally infected with a washed suspension of L. major amastigotes prepared from infected hamster skin tissue. The sand flies were permitted to feed on the suspension through a hamster cheek-pouch attached to a glass feeder warmed by a circulating water bath (Bastien 1990). The suspension was previously diluted with inactivated rabbit blood to give an estimated 3.5 x 105 viable amastigotes per 1 ml. Each fly with a full blood meal was estimated to have ingested approximately 100 parasites. Fed flies were maintained in a 20 x 20 x 20 cm insect cage, were provided with a solution of sucrose (50:50 v/v), and kept at 25°C, 95% RH and 16:8 D:L period. A random sample of the infected flies was taken at day 7 post-infection to verify the presence of parasites in the dissected gut. The chosen fly showed a high number of parasites when dissected and observed under light microscopy (Table). At day 13, infected sand flies were fed on a healthy anaesthetized hamster. In about 10 min, 4 flies were observed to have taken blood and were separated to be dissected. This practice was repeated at days 14, 18, 20 and 21 post-infection using new fresh hamsters to feed the remaining infected sand flies. In all cases the time of the feeding and the amount of blood taken were recorded (Table). The sand flies were then dissected-out and examined for infection in the SV, TMG and abdominal mid-gut (AMG). To analyze the morphology of the parasites in the infected sand flies, each part of the alimentary canal was teased apart on a glass slide, allowed to dry, fixed with methanol and stained with Giemsa's stain at 10% in phosphate buffer to observe for parasites. RESULTS From the 68 females of Ph. Papatasi primarily infected with L. major, 15 of them were able to survive oviposition. From these, 1 specimen was dissected at day 7 to verify the infection (Table). Nine out of the remaining 14 (64%) specimens, which were dissected after receiving a second blood meal on healthy hamsters, exhibited parasites in the alimentary canal. Different forms of Leishmania-parasite were seen in the gut of the sand flies at 13 to 21 days of infection. These included typical promastigotes (nectomonads and haptomonads), paramastigotes, metacyclic promastigotes and, in some samples, a large number of amastigote-like forms were also detected. The latter form was observed in 5 of the 9 infected flies (55%). In 2 of them (40%) this form was detected in the SV at days 14 and 18 post-infection; in 2 more flies (40%) similar forms were observed in the TMG at 13 and 21 days of infection, and in 1 more specimens (20%), the amastigote-like forms were detected in both the SV and the TMG (Table). These form were always found in groups and in some preparations they were distinguished together with a few typical promastigotes and/or paramastigotes. Fig. 1A, shows the quantity of amastigote-like forms of L. major detected in the SV of experimentally infected Ph. papatasi. A stained smear from a hamster lesion is included for comparison (Fig.1B). The comparison revealed a noticeable similarity in shape of the parasites from the two different sources. In no case were rounded forms seen in the AMG. DISCUSSION Leishmania amastigote-forms are normally found intracellularly within the macrophages of susceptible vertebrate hosts and have not been previously reported in any phlebotomine species as part of the parasite life cycle in the sand fly vector. They neither have been detected in the foregut of the infected sand flies, nor in the mass of parasites that colonize the anterior TMG, the SV or the esophagus after its luxurious development in the mid-gut. Furthermore, they appear to have gone unnoticed by other researchers using the pair Ph. papatasi-L. major (Warburg & Schlein 1986), or New World sand fly-Leishmania pairs (Nieves & Pimenta 2000). However, in the present work an impressive amount of massive, homogeneous amastigote-like forms were detected in the SV and the TMG of specimens of Ph. papatasi experimentally infected with L. major. The amastigote-like forms were obtained from Ph. papatasi 2 to 3 weeks post-infection and immediately after a second blood meal was taken from healthy hamsters. Two main factors are suspected to explain the presence of such forms in the infected flies. One may be the relatively low pH in the TMG (Tang et al. 1998), which induce promastigote transformation into rounded, amastigote forms (Bates & Tetley 1993). The other suspected factor appears to be the temperature of the blood directly ingested by the flies, from the vertebrate host which was approximately 5°C higher than that of the infected sand flies. Supporting evidence comes from experiments under in vitro conditions, in which axenic culture of New World Leishmania promastigotes were exposed to mammalian temperatures (33-37°C) resulting in morphological changes similar to those that occurred during transformation of promastigotes into amastigotes inside mammalian macrophages (Zilberstein & Shapira 1994, Valencia-Pacheco et al. 1998). The authors reported that promastigotes assumed a rounded form and lost their mobility because the flagellum shortened to the point where it no longer protruded from the flagellar pocket. In view of the similarity of the morphological forms obtained in the present work, we suspect that the same phenomenon might have occurred in the SV of L. major infected Ph. papatasi used in our experiments. Several arguments may be used to explain the presence of amastigote-like forms in the experimental model used, as well as to justify its possible incrimination as infective forms for the vertebrate. On one hand, it is known that the mechanism of entry by the Leishmania infective form from the infected sand fly into the mammalian host does not appear to contribute actively to the invasion process, rather relies on the phagocytic activity of the macrophage (Handman 2000). On the other hand, it has recently been stated that in the transition from the sand fly to the mammalian host, the promastigotes face two major environmental changes, a temperature shift to 35-37°C and a pH shift to around pH 5 (Antoine et al. 1998). The authors stated that the organisms detect the change in the new environment and transform into the obligatory intracellular amastigote with loss of flagellum and reduction in size. Considering this statement, in the context of our own findings, it is possible to suggest that the transition of promastigote to the amastigote-like form occurs in the sand fly itself during the act of feeding, as a consequence of the higher temperature of the uptaken blood from the host, and/or the low pH persisting in the TMG. From the biological point of view, this fact appears to be an advantage that may assure the deposition of the Leishmania infective forms on the vertebrate in a such a manner easily phagocytized by the macrophage or other antigen presenting cells such as the dendritic cells. Despite the suggestion that the effect of pH and temperature are factors that induce transition of promastigote into amastigote-like forms in the sand fly, details of how this phenomenon occurs are not yet understood and further investigation remains to discover unknown factors that, in addition to the changes of pH and temperature, are involved in this biological process. An additional argument to consider in regard to the L. major amastigote-like forms detected in Ph. papatasi that may actually be the infective forms to the vertebrate, is the necessity of such forms to be previously adapted to the hostile pH of the intracellular environment of the parasitophorous vacuole, as required for the amastigotes whose metabolism is adapted to an acidic pH (Glaser et al. 1988). This fact is supported by the results of Handman and Curtis (1982) who demonstrated that the membrane proteins of amastigotes are more resistant to proteolysis than promastigote membrane proteins. This also support Sacks (1989) opinion that metacyclogenesis is a preadaptation to survival within the vertebrate host, and those of Lawyer et al. (1990) who stated that the presence of the infective forms coincident with the host's search for another blood meal is a key factor in Leishmania transmission. Lawyer et al. (1990), after studying the development of L. major in Ph. duboscqi, stated that the presence of apparent intermediate forms between haptomonads and paramastigotes suggested that the latter is derived from the former. If this is true, it is possible to consider that the amastigote-like forms detected in the TMG and the SV of L. major-infected Ph. papatasi should derive from the observed paramastigotes, after a second blood meal was taken by the infected sand flies. To our knowledge, the development of massive, homogeneous Leishmania amastigote-like forms in the SV and TMG as observed in Ph. papatasi 13-21 days post-infection, and after a second blood meal, has not been previously reported. The relationships of these forms in the transmission to the vertebrate is not fully understood and requires further investigation. However, if the parasites from the SV can indeed be regurgitated and deposited on the skin of the vertebrate by the infected sand fly during feeding as suggested by Killick-Kendrick (1979), then it would be a highly effective means of transmission if the parasite population were to be comprised of forms directly preceding the amastigote form. This particular parasitic form, therefore, should easily be ingested by macrophages or another antigen presenting cells. Finally, the fact that 55% of the infected sand flies studied here presented the amastigote-like forms, appears to demonstrate that the present observations are not a chance happening. Indeed, the circumstance of finding the here-reported forms in the SV, TMG and in both sites of the alimentary canal in a range from 20-40% of the infected sand flies that received a second blood meal 2-3 weeks post-infection, seems to suggest that amastigote-like forms may play a role in the life cycle of Leishmania. ACKNOWLEDGEMENTS To Prof. I Woodward and Miss Kristen Bunch for carefully correction of the manuscript. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03107t1.jpg] [oc03107f1.jpg] |
| |||||||||

{kind=link}
{kind=link}