 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
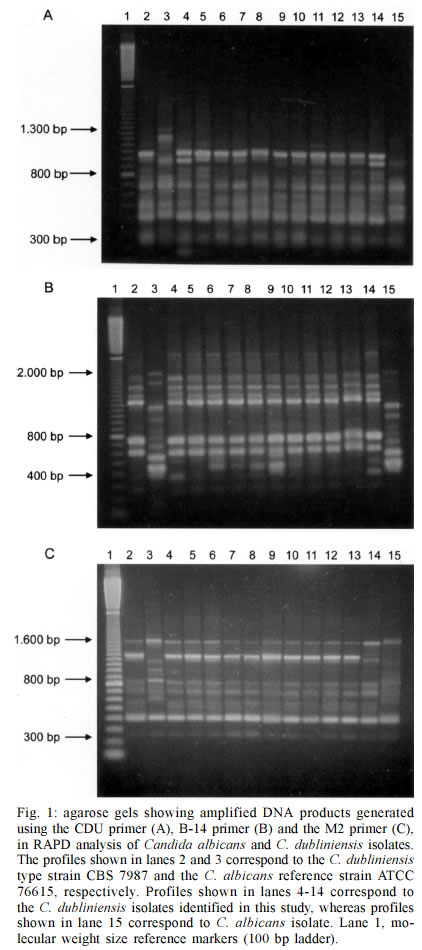
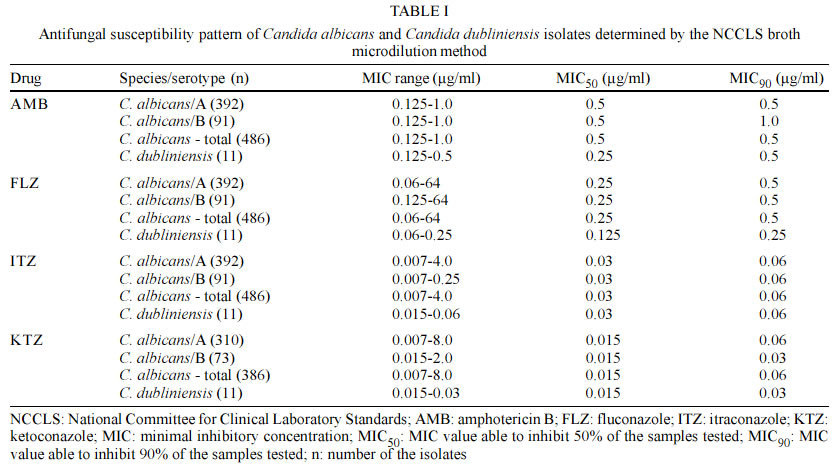
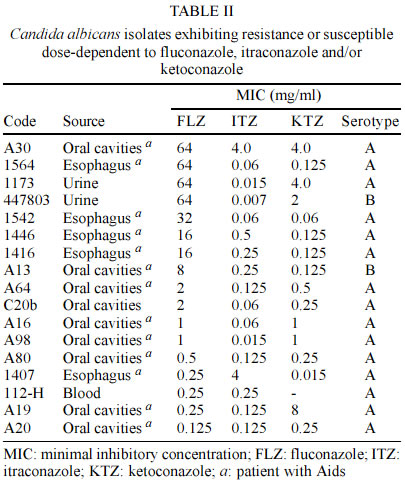
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 4, June, 2003, pp. 533-538 Candida dubliniensis Identification in Brazilian Yeast Stock Collection Priscilla de Laet Sant'Ana Mariano, Eveline Pípolo Milan/*, Daniel Archimedes da Matta, Arnaldo Lopes Colombo/+ Laboratório Especial de
Micologia, Disciplina de Doenças Infecciosas e Parasitárias,
Universidade Federal de São Paulo, Rua Botucatu 740, 04023-062 São
Paulo, SP, Brasil *Departamento de Microbiologia e Parasitologia, Universidade
Federal do Rio Grande do Norte, Natal, RN, Brasil This study was sponsored by Fundação de Amparo a Pesquisa do Estado de São Paulo, grants 99/ 07182-8 and 99/07134-3. Received 20 September 2002 Code Number: oc03116 We investigated the presence of Candida dubliniensis among isolates previously identified as Candida albicans and maintained in a yeast stock collection from 1994 to 2000. All isolates were serotyped and further evaluated for antifungal susceptibility profile. After doing a screening test for C. dubliniensis isolates based on the capability of colonies to grow at 42°C, its final identification was obtained by randomly amplified polymorphic DNA (RAPD) analysis using three different primers. A total of 46 out of 548 screened isolates did not exhibit growth at 42°C and were further genotyped by RAPD. Eleven isolates were identified as C. dubliniensis with RAPD analysis. Regarding serotypes, 81.5% of C. albicans and all C. dubliniensis isolates belonged to serotype A. Of note, 9 out of 11 C. dubliniensis isolates were obtained from patients with acquired immunodeficiency syndrome (Aids) and all of them were susceptible to azoles and amphotericin B. We found 17 (3%) C. albicans isolates that were dose-dependent susceptibility or resistant to azoles. In conclusion, we found a low rate of C. dubliniensis isolates among stock cultures of yeasts previously identified as C. albicans. Most of these isolates were recovered from oral samples of Aids patients and exhibited high susceptibility to amphotericin B and azoles. C. albicans serotype A susceptible to all antifungal drugs is the major phenotype found in our stock culture. Key words: Candida dubliniensis - oral candidiasis - Candida spp. - Aids Code Number: oc03116 Recently, Sullivan et al. (1995) described the novel species Candida dubliniensis, primarily associated with oral candidosis in human immunodeficiency virus (HIV) infected and acquired immunodeficiency syndrome (Aids) patients. C. dubliniensis shares many phenotypic characteristics with Candida albicans isolates and molecular methods are needed to distinguish both species (Sullivan et al. 1995, Coleman et al. 1997, Kirkpatrick et al. 1998, Sullivan & Coleman 1998). Probably, this new species has been present in the community for a long period of time and has been misidentified as C. albicans (Schorling et al. 2000). Definitive identification of C. dubliniensis is a problem in routine diagnostic laboratories and surveillance studies performed by reference laboratories are necessary to better understand the epidemiology of this new species. There are currently a wide variety of molecular techniques capable of identifying C. dubliniensis, including: DNA fingerprinting techniques using restriction endonuclease (REA) analysis, pulsed field gel electrophoresis-based methods, DNA fingerprinting using specific probes and polymerase chain reaction (PCR) based methods. Unfortunately, these methods are labor-intensive, expensive, and not readily applicable to routine use for identification purposes in most clinical microbiology diagnostic laboratories (Mannarelli & Kurtzman 1998, Kurzai et al. 1999, Sullivan et al. 1999, Park et al. 2000). In Brazil and South America there are a small number of reports addressing the epidemiology of this emergent species. The aims of this study were to identify C. dubliniensis strains among isolates from a large yeast culture collection including strains that were previously identified as C. albicans and to evaluate the serotypes and susceptibility profiles of isolates of both species. MATERIALS AND METHODS Samples - The isolates tested were selected among strains previously identified as C. albicans and maintained at the yeast stock collection of the Special Mycology Laboratory, Universidade Federal de São Paulo, between 1994 and 2000. The samples had been stored at -70°C in 10% glycerol, for different periods of time. We included strains isolated from different body fluids of HIV positive and negative patients as follow: blood, urine, esophageal and oropharyngeal samples. C. albicans identification - All isolates were inoculated on CHROMagar Candida medium (CHROMagar Microbiology, Paris, France) and incubated at 30oC for up to 5 days with daily examination looking for light green colonies (typical of C. albicans) and dark green colonies (typical of C. dubliniensis). Representative light green and/or dark green colonies were further purified by subculture on Sabouraud dextrose agar (SDA, Difco Laboratories, Detroit, MI) and were tested for their ability to produce chlamydoconidia on corn meal Tween 80 agar (Kirkpatrick et al. 1998, Odds et al. 1998). Serotyping assay - All strains were prepared for slide agglutination serotyping with serodiagnostic reagent number 6 from Candida Check kit (Iatron Laboratories, Inc., Higashi-Kanda, Chiyoda, Tokio, Japan), as suggested in the package insert. C. albicans cultures were incubated on Sabouraud-dextrose agar plates for 48 h at 25oC. Small amounts of specimen yeast cells were inoculated onto a Candida Check test tray and approximately 0.05 ml of specific serum (number 6) was added for testing and physiological saline was added as control. The glass test tray was stirred for about 1-2 min. A positive agglutination reaction was interpreted by the visualization of aggregates considered to be indicative of C. albicans serotype A. Negative results in the agglutination reaction with reagent number 6 were considered to be indicative of C. albicans serotype B (Sullivan et al. 1995). C. dubliniensis screening test - Following isolation on CHROMagar medium, a small portion of a single colony of each isolate was streaked for subculture over the surface on SDA agar plate and incubated at 42oC as previously described. All isolates that grew poorly or did not grow at all when incubated at 42oC were further genotyped by RAPD assays (Gales et al. 1998, Pinjon et al. 1998). RAPD analysis - Template DNA for RAPD analysis was prepared from yeast cells grown in 2 ml of YPD medium for 16 h at 30oC with agitation (220 r.p.m.) or until a density of 2 x 108 cells/ml be achieved. DNA was extracted using a rapid, small-scale isolation protocol described previously (Wash et al. 1994). RNA was removed by treatment with RNase A (Amersham Pharmacia Biotech, Piscataway, NJ, USA) for 1 h at 37oC. DNA concentration and purity were determined by optical density at 260 nm and ratio O.D. 260 nm/280 nm determinations, respectively. RAPD tests were also performed with template DNA from reference strains of C. albicans (ATCC 76615) and C. dubliniensis (CBS 7987). RAPD analysis was performed as described previously with the oligonucleotide primer CDU (5' GCG ATC CCC A 3') (Sullivan et al. 1995), M2 (5' CTT GAT TGC C 3') and the oligonucleotide primer B-14 (5' GAT CAA GTC C 3'), the latter previously reported by Bauer et al. (1993). Aliquots of 40 ng of total DNA were added to a 25 µl reaction containing 2.5 µl of a 10 x PCR buffer (100 mM Tris-HCl, pH 8.3, 500 mM KCl, 3.5 mM MgCl2), 4 µl of dNTP mix (1.25 mM each dNTP), 0.4 µM of primer, 0.5% (vol/vol) Tween 20 and 1.0 unit of Taq DNA polymerase (Amersham Pharmacia Biotech). Cycle conditions were 94oC for 1 min, 40oC for 1 min, 72oC for 2 min, for 45 cycles and a final extension at 72oC for 10 min, in a Perkin-Elmer DNA thermocycler (Gene Amp PCR System 9600). RAPD products were separated by electrophoresis for 3 h at 100 V in 1.5% (wt/vol) agarose gels using Tris-Acetate-EDTA (TAE) buffer. Following electrophoresis, gels were stained with a solution containing 0.5 µg/ml ethidium bromide for 10 min and destained twice for 15 min in 300 ml of distilled water. The gels were observed on UV transilluminator and photographed. RAPD profiles were defined with visual analysis, comparing the bands derived of the reference strains and the samples testing. Susceptibility testing - All isolates were tested for susceptibility to amphotericin B, fluconazole, itraconazole and ketoconazole by using a broth microdilution assay performed in accordance with the NCCLS M27-A standard (NCCLS 1997). Reference grade powders of fluconazole (Pfizer Inc., New York, NY, USA), itraconazole (Janssen Pharmaceutical, Titusville, New Jersey, NY, USA), ketoconazole (Janssen Pharmaceutical) and amphotericin B (Sigma Chemical Corporation, Saint Louis, MO, USA) were used to obtain final drug dilutions ranging from 0.125 to 64 µg/l, 0.007 to 4 µg/ml, 0.015 to 8 µg/ml and 0.03 to 16 µg/ml, respectively. Briefly, broth microdilution testing was performed in sterile, flat-bottom 96-well microplates (Costar, Corning Incorporated, New York, USA), with RPMI-1640 (Angus Buffers & Biochemicals, Niagara Falls, NY, USA) with L-glutamine, without bicarbonate, and buffered with MOPS at pH 7.0. The microplates containing the double of the final concentration of azoles were made in advance and stored at -70oC for no more than three weeks until use. On the test day, the inoculum suspension had a turbidity adjusted by the spectrophotometer to that produced by a 0.5 McFarland standard at 530 nm wavelength. A volume of 100 µl of the adjusted inoculum suspension was dispensed in each well resulting in the desired final drug concentration and inoculum size between 0.5 and 2.5 X 103 cells/ml. The plates were incubated at 35oC for 48 h. Quality control strain (C. parapsilosis ATCC 22019) was included on each day of the assay to check the accuracy of the drug dilutions and the reproducibility of the results. The Minimal Inhibitory Concentration (MIC) of amphotericin B was defined as the lowest concentration able to inhibit any visual growth. The MIC of azoles was the lowest concentration that exhibited prominent reduction in the turbidity, when compared with the growth control (drug-free) well (NCCLS 1997). Breakpoint definitions for fluconazole and itraconazole MICs were those proposed by the NCCLS (1997). Due to the lack of consensual definitions of breakpoints for ketoconazole and amphotericin B MICs, arbitrary values were established, following those suggested in previous studies (Rodriguez-Tudela et al. 1995, St-Germain et al. 1995, Milan et al. 1998). RESULTS A total of 548 isolates previously identified as C. albicans on the bases of phenotypic characterization were considered for analysis. Among the 548 samples tested on CHROMagar Candida medium, only 10 exhibited a dark green colony color while the others exhibited a common light green colony color. All of them produced chlamydoconidia on corn meal Tween 80 agar. The clinical sources of the isolates evaluated were: blood (100), urine (86), esophageal (97), and oropharyngeal samples (265). All blood and urine isolates were obtained from HIV negative critical care patients. All esophageal samples and 50% of the oropharyngeal isolates were obtained from HIV positive patients. After doing the temperature growth test, 46 isolates grew very slowly or not at all in these conditions and were further subjected to the RAPD analysis using three different primers. Based on RAPD findings, we concluded that 11 isolates exhibited the same profile of the CBS 7987 C. dubliniensis control organism. Among these 11 C. dubliniensis isolates found, only 2 exhibited a typical dark green colony color on CHROMagar Candida medium. Fig. 1 illustrates the RAPD banding patterns of the 11 clinical isolates obtained with RAPD using the 3 primers tested. Using the primer CDU, C. dubliniensis and C. albicans isolates yielded RAPD profiles with 7 and 6 strong bands, respectively, with molecular sizes ranging from 300 to 1300 bp. (Fig. 1A). The RAPD assay with primer B-14 yielded 6 strong bands to C. dubliniensis and 5 to C. albicans, with molecular sizes ranging from 400 to 2000 bp (Fig. 1B). Finally, using the primer M-2 we got 8 strong bands with C. dubliniensis and 5 with C. abicans isolates, with molecular sizes ranging from 300 to 1600 bp (Fig. 1C). The other 35 isolates produced RAPD profiles very similar to that obtained with the C. albicans reference strain ATCC 76615. All C. dubliniensis isolates belonged to serotype A. C. albicans isolates serotype A responded for 81.5% of all C. albicans strains. The distribution of C. albicans serotypes was very similar among isolates obtained from different clinical sources. The 11 C. dubliniensis isolates exhibited MIC values ranging from 0.125 to 0.5 for amphotericin B, 0.06 to 0.25 µg/ml for fluconazole, 0.015 to 0.06 µg/ml for itraconazole and 0.015 to 0.03 µg/ml for ketoconazole. These data indicated that all of the C. dubliniensis isolates were susceptible to the 4 antifungal drugs tested. The range of the MIC values obtained with C. albicans isolates was larger than that obtained with C. dubliniensis isolates, as illustrated in Table I. It was possible to identify 17 C. albicans isolates resistant and/or with susceptibility dose dependent (SDD) to azoles, most of them (76%) obtained from Aids patients. The MIC values exhibited by the azoles SDD/resistant isolates were presented in Table II. DISCUSSION In the present series we found a low prevalence rate of C. dubliniensis (2%) among 548 yeast isolates obtained from a stock culture collection consisting of isolates that had been previously identified as C. albicans. The evaluated isolates were collected from different clinical sources of HIV positive and negative patients. As expected, the 11 C. dubliniensis isolates were obtained from oropharyngeal samples and 9 of them were cultured from HIV positive patients. C. dubliniensis isolates have been mostly reported among Aids patients. The most extensive reported data of the incidence of C. dubliniensis in HIV infected and Aids patients comes from Ireland with a frequency about 18-32% (Coleman et al. 1997, Sullivan et al. 1999). Studies conducted in the United States have reported oral incidence rates for C. dubliniensis in HIV infected individuals ranging from 11.1 to 17.5% (Kirkpatrick et al. 1998, Meiller et al. 1999). In South America, the prevalence of C. dubliniensis isolates seems to be lower than that exhibited in north hemisphere countries. A recent prospective and multicenter study conducted by Milan et al. (2001), including the evaluation of 108 Brazilian Aids patients with oropharyngeal candidiasis, found that only 3 of them were harboring C. dubliniensis at the time they were sampled. In accordance with the mentioned findings by Milan et al. (2001), other authors from Latin American medical centers have reported only limited number of Aids patients who were colonized and/or infected by C. dubliniensis (Sano et al. 2000, Alves et al. 2001). The prevalence rate of C. dubliniensis found in our yeast stock collection was similar to that reported by similar studies performed by other investigators. Odds et al. (1998), using molecular typing for yeast identification, evaluated 2,589 isolates from stock cultures of yeasts originally identified as C. albicans and found that 2.1% of them were correctly identified as C. dubliniensis. In a similar study performed by Jabra-Rizk et al. (2000) the prevalence of C. dubliniensis identification was 1.2% among the 1,251 isolates previously identified as C. albicans. In the present study we found two C. dubliniensis isolates from oropharyngeal cultures obtained from HIV negative health carriers. These findings are in accordance with previous report of Coleman et al. (1997) that reported 3% of prevalence of this new species in health carriers from Ireland. Isolation of C. dubliniensis from clinical sources other than oropharyngeal samples obtained from HIV negative patients have been reported with increasing frequency. C. dubliniensis isolates have been isolated from different clinical sources, including blood, urine, vaginal and feces from diabetic patients, surgical patients, and patients submitted to organ transplantation (Meis et al. 1999, Brandt et al. 2000, Willis et al. 2000). Several methods for screening of C. dubliniensis have been reported, including culture on CHROMagar Candida, evaluation of their ability to grow at 42 or 45°C, evaluation of b-D-glucosidase activity as well as their carbohydrate assimilation pattern (Sullivan et al. 1995, 1997, Schoofs et al. 1997, Pinjon et al. 1998, Salkin et al. 1998). Of note, as mentioned by other investigators, most of our cultures of C. dubliniensis that had been stocked by long periods of time and did not exhibited the typical dark green colony color on CHROMagar Candida medium when incubated up to 5 days (Schoofs et al. 1997, Sullivan et al. 1999). In our study, the inability of the organism to grow at high temperature (42°C) was selected as a screening test for C. dubliniensis because it is inexpensive and easy to perform, generates results after 48 h incubation with a clear reading criteria and has been successfully used by different authors (Kirkpatrick et al. 1998, Tintelnot et al. 2000). In the present study, the final identification of C. dubliniensis was obtained by RAPD analysis, a method standardized in our laboratory that has been used by several investigators as a molecular tool for the identification of microorganisms (Sullivan et al. 1995, Coleman et al. 1997, Melo et al. 1998, Park et al. 2000, Milan et al. 2001). It is noteworthy that the 3 different primers we used for RAPD analysis exhibited several high density bands that allowed a clear visual interpretation of the DNA fingerprinting exhibited by C. dubliniensis and C. albicans isolates. All isolates of C. dubliniensis we found belonged to serotype A. The literature reports that C. dubliniensis isolates are able only to express serotype A phenotype of C. albicans (Schoofs et al. 1997, Sullivan & Coleman 1998). Regarding to the antifungal susceptibility profile of the isolates, all them were susceptible to azoles and amphotericin B. Pfaller et al. (1999a), after evaluating a total of 71 C. dubliniensis isolates, found that most of them are highly susceptible to amphotericin B and azoles. In Brazil, C. albicans serotype A susceptible to all antifungal drugs seems to be the most prevalent phenotype of C. albicans isolates, regardless of the origin of the sample. We found a frequency between 75.6% and 88.4% of serotype A among isolates obtained from HIV negative and positive, including all different clinical sources. This finding is in accordance with previous publications (Pires et al. 1996, Sant'Ana et al. 2002). Despite the high susceptibility to azoles exhibited by the 486 C. albicans isolates, we found 17 (3.5%) strains exhibiting susceptibility dose dependent status or resistance to azoles. It is noteworthy that 13 of the mentioned less susceptible isolates were recovered from Aids patients. C. albicans isolates resistant to fluconazole has been mainly reported among Aids patients with history of a long period of exposure to azoles (Persons et al. 1991, Millon et al. 1994, Barchiesi et al. 1996). In conclusion, the clinical sources of C. dubliniensis in Brazil appears to be similar to that exhibited by isolates evaluated in other series of yeast stock cultures from Europe and United States. Most of the isolates were obtained from oral cavities of HIV positive and Aids patients. C. albicans serotype A susceptible to all antifungal drugs was the major phenotype of C. albicans isolates stocked at our yeast collection, regardless of the clinical source of the strains. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03116t1.jpg] [oc03116t2.jpg] [oc03116f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}