 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 4, June, 2003, pp. 565-568 8-Methoxy-naphtho[2,3-b]thiophen-4,9-quinone, a Non-competitive Inhibitor of Trypanothione Reductase Carlos L Zani/+, Alan H Fairlamb* Laboratório de Química
de Produtos Naturais, Centro de Pesquisas René Rachou-Fiocruz, Av.
Augusto de Lima 1715, 30190-002 Belo Horizonte, MG, Brasil *Division of Biological
Chemistry & Molecular Microbiology, School of Life Sciences, The Wellcome
Trust Biocentre, University of Dundee, Dundee, UK Sponsorship Fiocruz/CNPq/Pronex/The British Council/Wellcome Trust Received 20 December 2002 Code Number: oc03122 The enzyme trypanothione reductase is a recognised drug target in trypanosomatids and has been used in the search of new compounds with potential activity against diseases such as leishmaniasis, Chagas disease and African trypanosomiasis. 8-Methoxy-naphtho [2,3-b] thiophen-4,9-quinone was selected in a screening of natural and synthetic compounds using an in vitro assay with the recombinant enzyme from Trypanosoma cruzi. Its mode of inhibition fits a non-competitive model with respect to the substrate (trypanothione) and to the co-factor (NADPH), with Ki-values of 5 and 3.6 µM, respectively. When tested against human glutathione reductase, this compound did not display any significant inhibition at 100 µM, indicating a good selectivity against the parasite enzyme. Key words: trypanothione reductase - Trypanosoma cruzi - Leishmania - enzyme inhibitors - naphthoquinone Protozoa of the order Kinetoplastida occur in vast areas of tropical and subtropical regions of the world. Many of them are able to infect humans and cause diseases such as Chagas disease (American trypanosomiasis) caused by Trypanosoma cruzi, sleeping sickness (African trypanosomiasis) caused by Trypanosoma brucei gambiense or rhodesiense and leishmaniasis caused by various Leishmania spp. (http://www.who.int/tdr/diseases/default.htm). Current chemotherapeutic agents are either not effective for all stages of these diseases, present serious side effects or require a complicated administration regimen. These parasites share a defence mechanism against oxidative stress based on the dithiol trypanothione [N1,N8-bis(glutathionyl)-spermidine] and associated enzymes, specially trypanothione reductase (TR) (Fairlamb et al. 1985, Fairlamb & Cerami 1992, Flohe et al. 1999). This enzyme was shown to be essential for these parasites survival and infectivity and is recognised as an important target for the development of new drugs against these diseases (Fairlamb 1999, Iribarne et al. 2002). A lot of effort has been devoted to find TR inhibitors. Using either rational or empiric approaches, different classes of compounds, including peptides and peptoid, substituted polyamines, quinacrine and acridine analogues, 2-aminodiphenylsulfides, phenothiazine derivatives, substituted piperazines and nitrofuran derivatives, were studied for their inhibitory potential (see reviews by Werbovetz 2000, Augustyns et al. 2001, Schmidt & Krauth-Siegel 2002). Investigation of natural products, although not so extensive, disclosed interesting inhibitors such as ajoene from garlic (Gallwitz et al. 1999), bisbenzyl-isoquinoline alkaloids (Fournet et al. 1998, 2000), and polyamines such as lunarine (Bond et al. 1999) and kukoamine (Ponasik et al. 1995). After the initial work of Henderson and co-workers (1988) naphthoquinone derivatives were further investigated for their action on TR (Jockers-Scherubl et al. 1989, Salmon-Chemin et al. 2000, 2001). In a previous work we reported the in vitro effect of several naphtho-thiophenquinone derivatives against epimastigote and trypomastigote forms of T. cruzi and the inhibition of the recombinant enzyme trypanothione reductase (Zani et al. 1997). We showed that quinone 8-Methoxy-naphtho[2,3-b]thiophen-4,9-quinone (TNQ2) was able to inhibit the enzyme activity by 87% at 100 µM after 30 min incubation. This compound was also one of the most active among 150 natural and synthetic compounds evaluated against TR (unpublished results). These findings stimulated us to carry out a more detailed investigation of the inhibition kinetics of this compound against TR and human glutathione reductase (hGR), the results of which are presented in this paper. MATERIALS AND METHODS 8-Methoxy-naphtho[2,3-b]thiophen-4,9-quinone - This compound was available from our previous work (Zani et al. 1997). Before use the material was recrystallized from MeOH-AcOEt to afford bright yellow needles with purity higher than 99.5%, as determined by reversed phase HPLC. A 100 mM stock solution was prepared in DMSO. This solution was diluted in the assay buffer to attain the desired concentrations in the assays. In vitro assay with T. cruzi TR - Recombinant T. cruzi TR was obtained as described by Borges et al. (1995). Enzyme activity was determined spectrophotometrically at 340 nm in 40 mM HEPES, pH 7.4, containing 1 mM EDTA, 200 µM NADPH, and 20 nM TR (assay volume 1 ml). Enzyme was pre-incubated at 27°C with 100 µM test compound for 5 min prior to addition of the substrate trypanothione disulphide (50 µM). Control assays contained an equivalent amount of DMSO. The results were expressed as percent reduction in the trypanothione reductase activity. In vitro assay with hGR - hGR was purified from human erythrocytes (Worthington & Rosemeyer 1974). Enzyme activity was determined under conditions similar to that used for TR, using hGR at 50 nM and glutathione at 100 µM. The stock solution of TNQ2 was diluted in the assay buffer to attain the desired concentrations. Control assays contained an equivalent amount of DMSO. The results were expressed as percent reduction in the hGR activity. Kinetic measurements - Initial
enzyme velocities (v0) were measured using a Shimadzu UV-2401
spectrometer equipped with a sample changer and controlled by a PC running
the kinetics software package. In experiments where the co-factor (NADPH)
was the variable substrate, the substrates (trypanothione disulphide in the
case of TR and glutathione disulphide in the case of hGR) were held constant
at 100 µM. When the disulphide substrate was varied, NADPH was 200 µM.
Kinetic parameters were obtained by non-linear fitting of v0 against
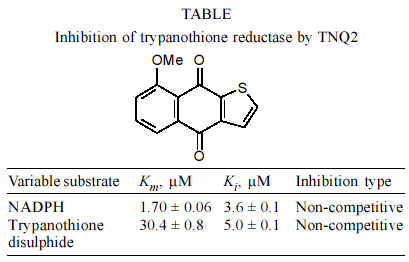
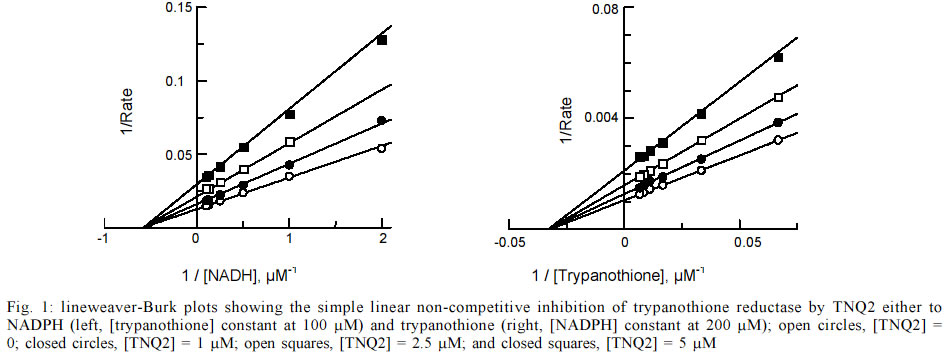
a range of substrate and inhibitor concentrations using GraFit software v.4.5. RESULTS AND DISCUSSION The naphthothiophenquinone TNQ2, available from our previous work (Zani et al. 1997), was re-crystallised from methanol-ethyl acetate to more than 99.5% purity, as assessed by HPLC analysis. A stock solution at 100 mM was prepared in DMSO and appropriate dilutions were prepared in the assay buffer. We initially checked that the incubation time of TNQ2 with TR does not affect its activity (data not shown) ensuring that the inhibition is not time dependent. Initial rates (v0) were then measured after 5 min incubation before the addition of the trypanothione with vigorous mixing. The decrease in the absorbance at 340 nm was measured and initial rates determined using the first 10-20 sec of data, in such way that the correlation coefficient to a straight line was greater than 0.95. Non-linear regression of the raw data was performed using GraFit to calculate the kinetic parameters Km and Ki (Table). The graphical results are presented in Fig. 1 as Lineweaver-Burk plots, showing a clear simple linear non-competitive profile with respect to both NADPH and the substrate trypanothione. TNQ2 was also tested on hGR, the enzyme with a similar function in the mammalian host. At 100 µM the activity of this enzyme was reduced by only 15% (data not shown), indicating a much lower activity against the host enzyme and thus a good degree of selectivity. In the case of a non-competitive inhibitor, the relative activity of TR can be calculated as follows:
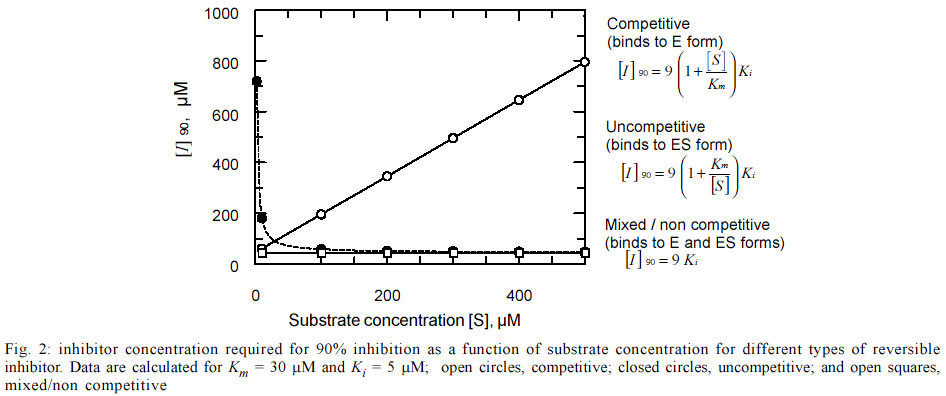
where vi and v0 are the inhibited and uninhibited rates for trypanothione reductase, respectively, Km is the Michaelis constant, [S] and [I] are concentrations of substrate and inhibitor, respectively and Ki the inhibitor constant. When vi/v0 = 0.1, it can be seen by rearranging the above equation that the concentration of inhibitor required for 90% inhibition ([I]90) is equal to 9Ki. Similar equations can be derived for competitive and uncompetitive inhibition (see Fig. 2). Since intracellular levels of reduced trypanothione are ~1 mM in T. cruzi (Ariyanayagam & Fairlamb 1997) the behaviour of these types of reversible inhibitor is presented as a function of substrate concentration up to 0.5 mM trypanothione disulphide. Note that, unlike competitive and uncompetitive inhibition, the [I]90 for non-competitive inhibition is independent of the substrate concentration. Thus, even though trypanothione disulphide levels would be expected to increase in cells due to the metabolic roles of trypanothione as an antioxidant and as an electron donor for reduction of ribonucleotides, the level of inhibition of trypanothione reductase would remain constant. Data from our previous work (Zani et al. 1997) shows that TNQ2 inhibited growth of epimastigotes with an IC50 of 14.3 µM. Assuming TNQ2 equilibrates across the membrane of the parasite, then, for [I] = 14.3 µM (50% inhibition of growth) and the Ki values given in the Table, TR would be inhibited by about 75%. In studies on conditional knockouts of trypanothione reductase in T. brucei, it was noted that growth completely ceased when enzyme levels fell to 95% of normal, but that lysis occurred only at even lower levels (Krieger et al. 2000) . Also, in L. donovani expressing a dominant-negative mutant form of trypanothione reductase, growth was unaffected by up to 85% reduction in activity, although sensitivity to oxidant stress was increased (Tovar et al. 1998). T. cruzi epimastigotes would appear to be slightly more sensitive than these organisms, bearing in mind the assumption made above about the intracellular levels of TNQ2 and assuming TNQ2 does not affect other cellular processes. This compound was only partially effective (27 ± 7% lysis) against the trypomastigote form of T. cruzi in blood at much higher concentrations (200 µM). This discrepancy could be due to factors such as: (a) the greatly reduced metabolic activity of T. cruzi at 4°C retarding the trypanocidal effect, (b) the reduced requirement for ribonucleotide reduction in non-dividing trypomastigote stages or (c) the difference in end point measurement (i.e. growth versus lysis). Since the most striking difference in the properties of TR and hGR is at their respective disulphide-binding sites, most compounds have been designed as competitive inhibitors. As discussed above, such inhibitors suffer from the disadvantage that their inhibitory effects are reversed by the accumulation of trypanothione disulphide. Irreversible inhibitors such as the antimonial drugs (Cunningham & Fairlamb 1995) or lunarine (Bond et al. 1999) or its analogues (Hamilton et al. 2003), or non-competitive inhibitors such as TNQ2 may offer a more fruitful approach. Work is in progress to identify the binding site for TNQ2 in order to increase potency and selectivity of this interesting lead. ACKNOWLEDGMENTS To Mr Ahilan Saravanamuthu for preparation of try-panothione reductase used in these studies. CLZ acknowledges the British Council-Fiocruz for a Travelling Fellowship to visit the UK. AHF is supported by the Wellcome Trust. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc03122t1.jpg] [oc03122f2.jpg] [oc03122f1.jpg] |
| |||||||||

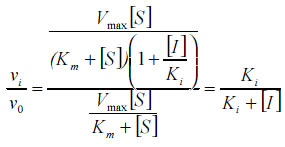
{kind=link}
{kind=link}
{kind=link}