 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
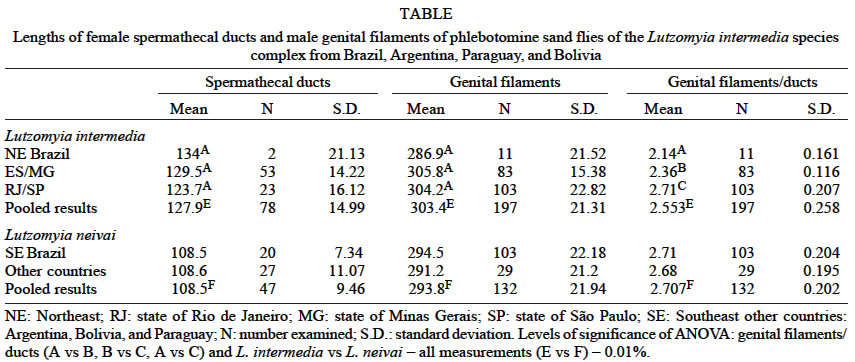
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 5, July, 2003, pp. 611-613 SHORT COMMUNICATION Correlation of Male Genital Filaments and Female Spermathecal Ducts in New World Sand Flies of the Lutzomyia intermedia Species Complex (Diptera: Psychodidae, Phlebotominae) Carlos Brisola Marcondes/+, Bruce Alexander* Departamento de Microbiologia e
Parasitologia, Centro de Ciências Biológicas, Universidade Federal
de Santa Catarina,
88040-900 Florianópolis, SC, Brasil *Centro de Pesquisas René Rachou-Fiocruz,
Belo Horizonte, MG, Brasil Received 13 December 2002 Code Number: oc03129 The lengths of the male genital filaments and female spermathecal ducts were measured in phlebotomine sand flies of the Lutzomyia intermedia species complex and the ratios between these characters calculated. Ratios for L. intermedia s. s. from Northeast vs Southeast Brazil (Espírito Santo and Minas Gerais), Espírito Santo/Minas Gerais vs Rio de Janeiro/São Paulo and L. intermedia vs L. neivai were significantly different at P < 0.1, 0.05 and 0.01 respectively when compared using ANOVA. The spermathecal ducts and genital filaments of L. intermedia were significantly longer than those of L. neivai (P < 0.01) and could be used to differentiate these species. The taxonomic and biological significance of these differences is discussed. Key words: Lutzomyia intermedia - Phlebotominae - Lutzomyia neivai - morphometry - reproduction Phlebotomine sand flies, particularly those of the New World genus Lutzomyia, exhibit considerable mor-phological variation with respect to the genitalia of both sexes. Unlike most Diptera, including the other members of the family Psychodidae, the organs of sperm transfer in phlebotomines present a bipartite condition (Downes 1968). The principal morphological characteristics used to distinguish between species are the external genitalia of the male, consisting of paired structures that grip the female during copula, and the internal genitalia of the female, consisting of a common duct branching to paired individual ducts terminating in spermathecae. The testes of the males are connected to a genital pump, which connects via the aedeagus to a pair of "genital filaments" (actually ducts). These filaments are the structures that penetrate the body of the female and deposit the spermatozoa, rather than the aedeagus as in most insects. Hertig (1949) was able to anaesthetise, clear and slide mount a copulating pair of Phlebotomus perfiliewi Parrot, showing the genital filaments of the male entering the genital ducts of the female, with their extremities almost reaching the spermathecae. The relationship between male genital filaments and spermathecal ducts has been studied in few other sand fly species. Forattini (1973) reviewed previous studies on copula in phlebotomine sand flies and illustrated two insects in copula of a species he referred to as "Psychodopygus intermedius"; these were later identified as L. neivai (Pinto) (Marcondes 1997). Ilango and Lane (2000) observed considerable variation among 26 Old World species with respect to the ratio between the lengths of the male genital filaments and female spermathecal ducts. The ratio between the length of male genital filaments and that of spermathecal ducts of their co-specific females may be important to the lock-and-key mechanism of the beetle Bambra invisibilis (Dybas & Dybas 1981). In view of the relative lack of information on this topic in New World species, the morphology of members of the Lutzomyia intermedia species complex was studied. This taxon includes L. intermedia (Lutz & Neiva) and L. neivai (Pinto), the latter resurrected by Marcondes (1996). The females can be easily differentiated by the spermathecae and by the lengths of the spermathecal ducts, besides the number of cibarial horizontal teeth. Some proportions between genital and extra-genital structures (Marcondes & Borges 2000) can differentiate males. The two species have rather similar ecological re-quirements, occurring in habitats modified by human activities as well as some preserved forest areas (Gomes & Galati 1989). However L. neivai occurs in colder, drier regions of Brazil than L. intermedia (Marcondes et al. 1998e) as well as Argentina, Paraguay and Bolivia (Marcondes et al. 1998b, e). In the Brazilian state of São Paulo, L. intermedia occurs between the eastern chain of mountains ("Serra do Mar") and the coast, while L. neivai has a more westerly distribution. The two species are sympatric at Pariqüera Açu (Marcondes 1996, 1998e). Although neither species meets all the requirements of a vector cited by Killick-Kendrick (1990) both L. intermedia and L. neivai have been incriminated as vectors of Leishmania. The genital ducts or filaments of 454 specimens (125 females and 329 males) of the L. intermedia species complex from Brazil, Argentina, Paraguay and Bolivia were measured during the present study, using an Olympus OSM- 221939 ocular. Insects were mounted using Canada balsam or NC (Nelson Cerqueira, Enecê in Portuguese), correcting the dimensions of the genital filaments according to Marcondes et al. (1998a). The insects were distributed among five groups, according to the species and region. The ratios between the means of the sum of the lengths of common and individual spermathecal ducts of females and the lengths of the genital filaments of males from the same region were obtained. Insects from localities at two distinct elevations in the state of Espírito Santo were also compared. Dimensions and ratios of the different regions were compared, analysing results by ANOVA, using Excel software. Mean lengths of spermathecal ducts and genital filaments (+ 1SD) of specimens of L. intermedia s.s. from three areas of Brazil (Northeast, Espírito Santo/Minas Gerais and Rio de Janeiro/São Paulo) and three other countries (Argentina, Paraguay, and Bolivia) are shown in the Table. The differences between the two species with respect to the lengths of the structures and genital filament/ spermathecal duct ratios were highly significant, as were some intraspecific differences between the ratios in L. intermedia from different regions. No significant differences were found either in the genital structures or in the ratios in the insects from the localities at different elevations; this agrees with the observations of Marcondes et al. (1999), who did not test the sum of the spermathecal ducts and the filament/duct ratio. Since in the present study insects from several localities were measured for each group, it would be useful to compare ratios from well-defined localities. The proportion between these lengths could therefore be used as a character to differentiate between these species (Marcondes 1996, Marcondes et al. 1998b, c, d, Marcondes & Borges 2000). Geographical variation with respect to this ratio should also be studied. Ilango and Lane (2000) noted that the common spermathecal duct of P. argentipes is very large and the male aedeagus correspondingly wide. Since females of L. intermedia have long and relatively large common ducts and those of L. neivai very short ones, it would be interesting to study the shape and dimensions of the male aedeagi and their relationship with the common ducts. The results of the present study demonstrate that the genital filaments are longer than the spermathecal ducts, as found for 25 of the 26 taxa examined by Ilango and Lane (2000). This greater length of genital filaments would allow sperm displacement by males (Ilango, 1995 apud Ilango & Lane 2000). These observations contradict the conclusion that lengths of genital filaments and those of spermathecal ducts are similar in sand flies, as proposed by Williams (1988). Large numbers of species may occur together within a relatively small area, such as the 35 recorded at a single collecting site in Panama by Chaniotis et al. (1972). There would be a considerable risk of interspecific couplings in a situation of this type if the insects did not have some method of recognising members of the same species. Although sex pheromones have been described for a few species (Hamilton et al. 1999) such isolating mechanisms are unknown for most sand flies. Nevertheless it is unlikely that the morphological diversity of the genitalia in Lutzomyia spp. simply acts as a physical barrier to hybridisation. Eberhard (1985) discussed the reasons why male copulatory structures in many groups of animals exhibit such variation in morphology. Two explications have been proposed, i.e., the "lock and key" and pleiotropy hypotheses. With respect to the first hypothesis, there are many groups of animals in which the complex genitalia of the females is unable to exclude the genitalia of males of other species. In pleiotropy, physical characteristics are effects of genes that code primarily for other characteristics, resulting in adaptations to the en-vironment. This idea, however, does not explain why these incidental effects occur principally in genitalia rather than in other organs. According to Eberhard (1985), two mechanisms proposed could be involved: competition among males and selection by females. In approximately 30% of the species of insects studied, the males carry out some type of behaviour apparently designed to stimulate the female. In phlebotomines these could include vibration of the wings during copula and mechanical stimulation of the female abdomen by tufts and other structures on the copulatory apparatus of the males, as seen in some butterflies (Lorkovic 1952). Large congregations of male sand flies at the bases of trees and other so-called diurnal resting sites probably represent courtship "leks" (Memmott 1991) where the males wait for the females to descend from the canopy to lay their eggs in the soil or leaf litter. Fertilisation of the eggs occurs shortly before oviposition and it would thus benefit a male sand fly to be the last to mate with a female. The females of many species of insects mate a second time before the sperm from the first insemination is exhausted. When this occurs, the spermatozoa of the second male preferentially fertilise the remaining eggs (Gwynne 1984). In damselflies, males remove the sperm of previous matings, the tip of the aedeagus being modified for this purpose (Waage 1984). In some species of phlebotomines, (including many members of the subgenus Nyssomyia Barrettto, to which both L. intermedia and L. neivai belong, the tips of the genital filaments also resemble spoons or hooks, their function being unknown. It would be interesting to determine whether these are also involved in sperm replacement. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc03129t1.jpg] |
| |||||||||

{kind=link}