 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
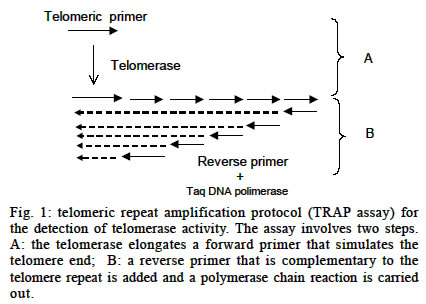
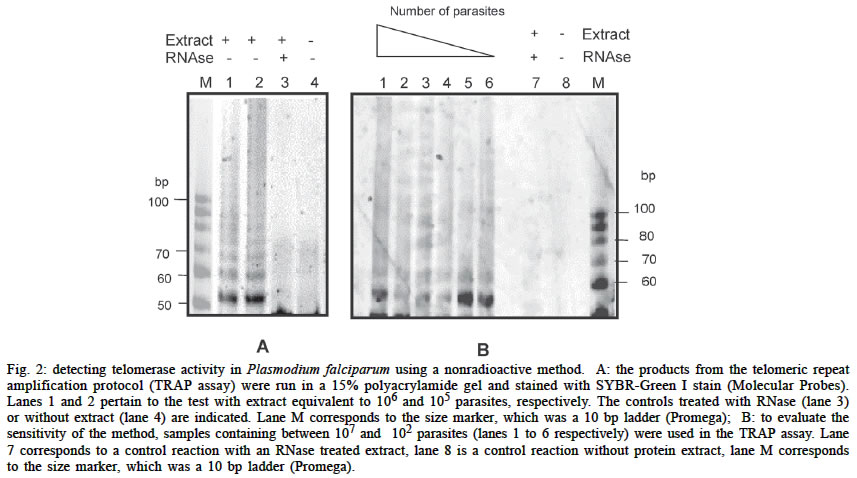
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 5, July, 2003, pp. 693-695 SHORT COMMUNICATION Detection of Telomerase Activity in Plasmodium falciparum Using a Nonradioactive Method Claudia C Rubiano, Moises Wasserman+ Laboratorio de Bioquímica,
Instituto Nacional de Salud, Bogotá, Colombia and Laboratorio de Investigaciones
Básicas en Bioquímica, Departamento de Química, Facultad
de Ciencias, Universidad Nacional de Colombia, Bogotá, Colombia This work was supported by the Instituto Nacional de Salud, by the Universidad Nacional de Colombia, project DINAIN 2010100198 and by the Instituto Colombiano para el Fomento de la Ciencia y la Tecnología Colciencias project 1101-05-11422. Received 24 January 2003 Code Number: oc03142 A simple, quick and sensitive method was used to detect telomerase activity in Plasmodium falciparum. The telomeric repeat amplification protocol (TRAP assay) was modified using electrophoresis and staining with SYBR-green I to detect telomerase activity in a range of 102 to 107 parasites. This might be a useful way to ascertain telomerase activity in different types of nontumor cells. Key words: Plasmodium - malaria - telomerase - assay - nonradioactive Telomerase is the enzyme responsible for compensating DNA loss at the ends of linear chromosomes in most eukaryotic cells (Greider & Blackburn 1985, Greider 1996). It is a reverse transcriptase with its own RNA, which it uses as a template to synthesize the telomeric repeats over again (Blackburn et al. 2000). Telomerase activity has been detected in 85-90% of human tumors and cell lines derived from tumors (Counter et al. 1994, Kim et al. 1994, Landberg et al. 1997) and it was proposed that its inhibition could detain growth, primarily in cells with high replication rates (Herbert et al. 1999, Liu 1999). The telomeric repeat amplification protocol (TRAP assay) (Kim et al. 1994) and several of its modified ver-sions have become the standard method for measuring telomerase activity in different types of cells. This assay involves elongation by telomerase of a primer that simulates the telomeric end, followed by amplification of the elongated products in a polymerase chain reaction (PCR) with a reverse primer complementary to the elongated telomeric repeats (see Fig. 1). Using the TRAP assay, telomerase activity has been detected in Plasmodium falciparum, which is the parasite responsible for the most severe form of malaria in human beings. This has been accomplished through radioactive labeling of the products obtained (Bottius et al. 1998, Sriwilaijareon et al. 2002) or by using the kit TRAP-eze (Oncor, Gaithersburg, MD) (Aldous et al. 1998). This kit uses primers that are not specific for P. falciparum, whose telomeric repeats differ from other eukaryotes. In the present study, telomerase activity in P. falciparum was detected using the TRAP assay with specific primers for Plasmodium and staining the products with SYBR-green I stain (Molecular Probes, Inc. Eugene, Oregon US), which is a highly sensitive method, already used to detect very low quantities of DNA (Karlsen et al. 1995, Schneeberger et al. 1995, Perkins. 2001, Aldea et al. 2002, Polanco et al. 2002). Parasites of the FCB-2 strain, cultivated according to the Trager and Jensen (1976) method, were used to prepare the protein extracts. Although not stringently syn-chronized, the majority of the parasites (80-90%) corresponded to trophozoites between 36-40 h old. The infected erythrocytes were isolated from the culture medium through centrifugation (2 500 x g, 10 min) and washed with HBS isotonic buffer (20 mM HEPES pH 7.4, 160 mM NaCl). The parasites were freed from the erythrocytes through lysis with 0.15% saponin in HBS (4ºC, 15 min) and centrifugation (15 000 x g, 15 min). The parasite pellet was washed two or three times with HBS (10 000 x g, 5 min) and 1/100 vol. of a mixture of protease inhibitors was added (1 mg/ml pepstatin A, 1 gm/ml leupeptin, 1 mg/ml aprotinium, 0.1 mg/ml phenantroline, and 1 mM Benzamidine-HCI) as well as an RNase inhibitor (40U RNase-out, BRL). It was then stored at -70ºC until extraction. The parasite pellet was resuspended in 200-400 µl of TMG buffer (10 mM Tris HCI pH 7.5, mM MgCl2, 10% glycerol, 10 mM β-mercaptoethanol), and 1/10 of 2% non-idet P40 in TMG buffer was added to lyse the parasites. The mixture was shaken for 30 min at 4ºC, then subjected to ultracentrifugation at 100 000 x g, at 4ºC for 1 h. The S100 supernatant or fraction was stored at -70ºC until its use. The protein concentration was determined by the Bradford method, using bovine serum albumin (BSA) in TMG buffer for the calibration curve. The PfTS-1 and PfCX primers described by Bottius et al. (1998), were used for the TRAP assay. They are specific for P. falciparum. The PfTS-1 primer (5' AATCCGTC GAGCAGAGTTCA 3'), which acts as a substrate for the telomerase, contains specific P. falciparum telomere sequences at the 3' end and the PfCX primer (5' GGCGCGT/GAAACCCTG/AAACCCTG/AAACCC 3'), which acts as a reverse primer for amplification, has 3 repeats complementary to the P. falciparum telomeric repeats and a GC clamp on the 5' end. The reactions were carried out on a total volume of 50 µl which contained TRAP buffer (20 mM Tris HCI pH 8.3, 1.5 mM Mg Cl 2, 0.005% Tween 20, 1 mM EGTA, 0.2 mg/ml BSA), 50 µM dNTPs, 5 U of RNase inhibitor (RNase-out, BRL), 0.1 µg of PfTS-1 primer and parasite protein extract (S100 fraction). This mixture was incubated for 1 h at 37ºC to allow the telomerase to elongate the primer, before adding 2.5 U of Taq DNA polymerase (Promega) and 0.1 µg of the PfCX primer to carry out the PCR reaction, which consisted of 35 denaturation cycles at 94ºC x 10 s, annealing at 55ºC x 30 s and extension at 72ºC x 1 min. An extract previously treated with 10 µg of RNase A for 30 min at 37ºC and a reaction without the protein extract were included as controls. Once the reactions were completed, a non-denaturant electrophoresis was done on 15% polyacrylamide gels in 0.5 x TBE buffer (44.5 mM Tris HCI, 44.5 mM boric acid, 1 mM EDTA pH 8.3). Subsequent to electrophoresis, the gels were immersed for 30 min in a solution of SYBR-green I stain (Molecular Probes), which was prepared in TBE 0.5 x buffer according to the manufacturer's recom-mendations, then visualized at UV (254 nm). The images were processed using the ONE-D-Scan (ScanalyticsÓ, CSPI division, US) program. The assays were considered positive when a ladder of products was observed with a difference of approximately 6 nt between the bands above 50 pb, as illustrated in Fig. 2A, where the reactions were done using protein extracts equivalent to 105 and 106 parasites. For an idea of the sensitivity of detection by this method, the TRAP assay was carried out with protein extract in amounts corresponding to a range of 102 to 107 parasites (see Fig. 2B). When the number of parasites is high (107), the signals diffuse and one obtains a smear more than a ladder of bands. Reducing the number of parasites gives a better definition of the signals, with the equivalent of 106 to 105 parasites being the optimum number for observing the signals. Below this number; that is from 104 to 102, the products can be observed, but with less intensity and definition. Although there have been some reports to date of electrophoretic detection of telomerase activity using nonradioactive methods (Fujita et al. 1998, Dalla Torre et al. 2000, Zhang et al. 2000), they have been applied to tumor cells or transformed cell lines where telomerase activity is significantly higher than in other types of cells. In this respect, the present study shows it is possible to evidence telomerase activity effectively in nontumor cells, using a nonradioactive method. Detecting telomerase activity is an important and necessary step in studies associated with this enzyme and its implications on cell proliferation. It is, therefore, convenient to have a simple and adequate method for this purpose. With nonradioactive detection, the TRAP assay becomes an easy-to-use tool; that is, a simple, quick and sensitive way to detect telomerase activity in protozoan parasites and possibly in other nontumor cells. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc03142f1.jpg] [oc03142f2.jpg] |
| |||||||||

{kind=link}
{kind=link}