 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
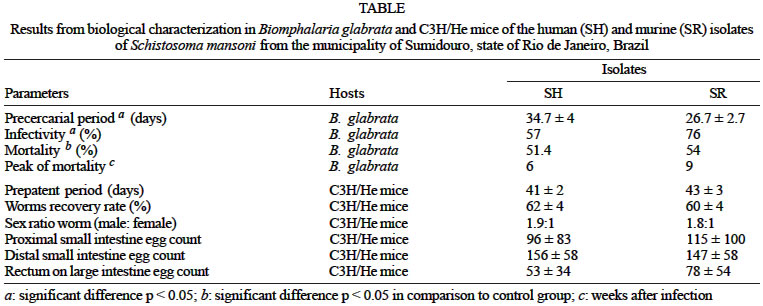
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 6, Sept, 2003, pp. 783-787 A Comparative Parasitologic Study on Biomphalaria glabrata Snail and C3H/He Mice Infected with Human and Murine Isolates of Schistosoma mansoni Derived from Sumidouro, Rio de Janeiro, Brazil Nilcéa Freire, Rosângela Rodrigues-Silva*, José Roberto Machado-Silva/+/++, Luís Rey**/++ Laboratório de Helmintologia
Romero Lascasas Porto, Disciplina de Parasitologia, Departamento de Patologia
e Laboratórios, Faculdade de Ciências Médicas, Uerj,
Av. Prof. Manuel de Abreu 444, 5º andar, 20550-031 Rio de Janeiro, RJ,
Brasil *Laboratório de Helmintos Parasitos de Vertebrados, Departamento
de Helmintologia **Departamento de Medicina Tropical, Instituto Oswaldo Cruz-Fiocruz,
Rio de Janeiro, RJ, Brasil Received 18 December 2002 Code Number: oc03159 Experiments were carried out to analyze the biological characteristics of two sympatric isolates of Schistosoma mansoni derived from humans and murines in a low endemic transmission area (Sumidouro county, state of Rio de Janeiro, Brazil). Sympatric reared-laboratory Biomphalaria glabrata and C3H/He mice were used as experimental hosts. Parameters assessed comprised: precercarial period, infectivity and mortality (snails), prepatent period, infectivity (percentage of cercariae maturation into adult worm) and intestinal egg count (mice). The murine isolate showed a shorter precercarial period and higher infectivity than human isolate (p < 0.05). This biological heterogenicity did not correspond to the vertebrate data because any biological parameter presented significant difference (p > 0.05). These data suggest that both isolates are local sub-populations, providing support for the hypotheses that in a same biotope mixed populations or sub-populations circulate among their main host (human beings) and/or rodent as an anfixenous infection. Key words: Schistosoma mansoni - human and murine isolates - host-parasite relationship - snail - C3H/He mice - Rio de Janeiro - Brazil Despite that humans are main hosts, rodents also harbor natural infection in both Africa and the Neotropical area (Combes 1990, Rey 1993, Alarcón de Noya et al. 1997, D'Andrea et al. 2000, Duplantier & Sène 2000). Thus, it is believed that rodents (alone or not) are able of participating on the schistosomiasis mansoni transmission depending on special ecological conditions (Alarcón de Noya et al. 1997). The coevolution of schistosomes and their freshwater snail hosts has given rise to selective pressures affecting the rate of either parasite or invertebrate host evolution. Thus, it has have demonstrated that this parasitic flatworm trematode exhibits a high degree of specificity to the snail host (Frandsen 1979). Furthermore, vertebrate hosts also behave as biological filters to parasites selecting phenotypes and genotypes within natural populations (Deane et al. 1984). It is widely known that Schistosoma mansoni geographical strains exhibit variability related to infectivity, prepatent period, fecal egg excretion, liver or intestinal egg counts and altered response to schistosomicidal drugs in rodent models (Kassim et al. 1979, Coelho et al. 1997, Incani et al. 2001, Yoshioka et al. 2002, Bonesso-Sabadini & Dias 2002). However, little attention has been paid to the biological characteristics of sympatric human and murine strains. Laboratory observations have shown that such populations from Paraíba do Sul River Valley (state of São Paulo) differ from each other because the murine isolate presents better adaptation to Biomphalaria tenagophila. Conversely, it was found that in a Lowland Region of the state of Maranhão, the human strain displayed a more suitable relationship to B. glabrata than a murine one (Bastos et al. 1982). Considerable information acquired from earlier studies carried out at Sumidouro (state of Rio de Janeiro) showed that in a same transmission area, either humans or rodents harbor schistosomiasis mansoni infection (Silva et al. 1992). Recently, it was demonstrated that cercariae deriving from sympatric murine and human isolates display only slight differences (Freire et al. 2002). Further studies of the parasitological characteristics of both sympatric isolates and epidemiological implications are presented and discussed. MATERIALS AND METHODS Schistosomes and definitive hosts - S. mansoni murine and human isolates were used in this study. The isolation methodology and conditions for the laboratory maintenance were detailed elsewhere (Machado-Silva et al. 1994). Briefly, the isolates of S. mansoni here studied were derived from a low endemic schistosomiasis mansoni transmission area: Pamparrão Valley, a rural area in the municipality of Sumidouro, about 200 km from the city of Rio de Janeiro, Brazil (D'Andrea et al. 2000). One of them was isolated from the feces of a natural S. mansoni-infected rodent Nectomys squamipes (2n = 56 chromosomes). Another one was recovered from the stools of human hosts who had acquired the infection in this same area. This last isolate was named SH while the first one was named SR. Snail infection - Sympatric laboratory-reared B. glabrata snails measuring 5 mm in shell diameter were individually exposed to 10 freshly hatched miracidia from each isolate: SR (n = 178) or SH (n = 182) in 10 ml of well water. Infection-free snails (n = 100) were used as a control group. The snails were kept in glass vials with de-chlorinated tap water and fed fresh lettuce (Lacttuca sativa) ad libitum (Freire et al. 2002). Eighteen days following snail exposure to hatched miracidia, and then weekly for up to 91 days, snails were exposed to artificial light. This procedure allowed the determination of the precercarial period (time required for miracidium penetration until first cercariae escape) (Frandsen 1979) and infectivity (percentage of infected snails shedding cercariae). The cercariae obtained from 40 B. glabrata were used to infect male or female seven-days-old C3H/He mice (n = 74). C3H/He mice were purchased from Centro de Criação de Animais de Laboratório, Instituto Oswaldo Cruz-Fiocruz, Brazil. For each isolate of S. mansoni parasites, 37 mice were individually infected with 50 cercariae, transcutaneously. Parasitological examination - In order to determine the prepatent period, animals were separated individually from the 28th day after exposure onward. For detection of fecal eggs, feces passed by each animal were collected and examined by the Kato-Katz technique. Two slides were analyzed per mouse for each examination. Adult worm recovery - Schistosomes were recovered from individual mice eight weeks post-infection by the conventional portal-hepatic perfusion. Worms were collected, counted and infectivity (percentage of cercariae maturation into adult worm) was calculated. Small and large intestine were removed and separated into three segments: proximal (between pylori and middle of the small intestine), distal (from this point until the end of the small intestine) and large intestine (rectum). From each segment, a fragment (1 cm long) was cut and crushed between two glass slides for egg count under brightfield microscopy, observations were expressed in egg/1 cm. Husbrandry - The rodents were housed conven-tionally in polypropylene cages (40 cm x 33 cm) with stainless steel screened covers. Animals received a well-fed conventional pellet mice diet (Nuvilab CR1, Colombo, Paraná, Brazil) and water ad libitum. The experiments reported here comply with the current laws regarding ethical procedures with investigated animals (S/GV-SOLAS 1985). Both isolates were studied in their first passage under laboratory conditions. Life span parameters were registered weekly. Statistical analysis - Experimental data were compared by means of T Student test, Z test, chi-square test and one-way analysis of variance (ANOVA). The statistical software BMDP-7M was used. P ≤ 0.05 was taken to represent significant difference. RESULTS Snail infection - The data obtained are shown in the Table. Significant differences (p < 0.05) were observed in the length of the precercarial period between SR isolate (26.7 ± 2.7) and SH isolate (34.7 ± 4), as well for infectivity 76% and 57.1% from SR isolate and SH isolate, respectively. The human isolate had an earlier peak of mortality (six weeks post-infection) than the murine one (nine weeks post-infection), although both isolates presented a similar mortality rate 51.4% and 54% from SH isolate and SR isolate, respectively. These rates differed significantly (p > 0.05) in comparison to control group (7%). Vertebrate infection - As appears in the Table, the mean prepatent period ranged 41 ± 2 days (SH isolate) and 43 ± 3 days (SR isolate). The infectivity rate was between 62 ± 4 % (SH) and 60 ± 4% (SR). The male/female sex ratio was biased toward males: 1.9:1 (SH isolate) and 1.8:1 (SR isolate). No significant differences (p > 0.05) were observed in those parameters. The mean egg count (egg/g feces) was similar for both isolates: 306 ± 61 and 340 ± 60 from SH isolate and SR isolate, respectively. The number of eggs trapped in the distal segment 156 ± 58 (SH isolate) and 147 ± 58 (SR isolate) was relatively greater than proximal 96 ± 83 (SH isolate) and 115 ± 100 (SR isolate). The number of eggs found in the large intestine 53 ± 34 (SH isolate) and 78 ± 54 (SR isolate) was significantly different (p < 0.05) from other two segments. No significant linear correlation was demonstrated between overall tissue egg count and the number of adult worms recovered from both isolates. DISCUSSION In this study it was shown that both isolates share several biological characteristics. Parasites are living organisms that are exposed to selective pressures forced by vertebrate and invertebrate hosts, or both. It is well known that geographic strains display several levels of adjustment to B. glabrata (Frandsen 1979, Incani 1993). The time required for the development of parasitic flatworms into snails, miracidia infectivity and snail mortality are biological indicators of the adjustment of the parasite into the intermediate host snail. Experimental studies have shown that the most compatible between this type of organisms are characterized by: development within the shortest time (Frandsen 1979), high infectivity (Zanotti-Magalhães et al. 1991), low mortality (Bastos et al. 1978) and great cercariae shedding (Incani 1993). Based on shorter precercarial period, higher infectivity and later peak of mortality, the murine isolate seems to be more adapted. This study provided interesting insight into the biology of schistosomes and arised some questions. Some researchers advocate that compatibility depends on the time lapsed between parasite-snail relationships. It is well established that the murine schistosomiasis occurred earlier than human infection (Combes 1990). In this context, older associations are less harmful (Bastos et al. 1982). In Brazil, several S. mansoni populations have been restricted to the entrance port, slave commerce and agricultural areas on the Brazilian coast (Rey 1993). From there, the endemic area gradually expanded northwards and southwards along with other agricultural crops (Grault et al. 1998). The Neotropical isolates have diverged from the original African population strains as observed by a better affinity to rodents. Meanwhile, when and how Brazilian autochthonous small mammals became infected still remains to be elicited. It could speculate that a paleoparasitology study could be carried out in animal (rodents) coprolites aiming to demonstrate S. mansoni eggs as recently evidenced in archeological material from France (Bouchet et al. 2002). The mouse model has yielded numerous results showing infraspecific variation in S. mansoni infection. Thus, it has been well documented that the mean pre-patent period is five-six weeks post infection (Kassim et al. 1979). Strains that have been maintained for a long period of time under laboratory conditions had a smaller prepatent period compared to newly isolated strains (Fallon et al. 1997, Incani et al. 2001, Yoshioka et al. 2002). In the present work, the murine isolate presented a faster precercarial period. However, no significant differences were found in relation to the prepatent period in mice. Other previous Brazilian investigations have demonstrated that isolates from patients with various clinical forms of schistosomiasis have a similar prepatent period (Costa & Katz 1982). Comparative studies have demonstrated that a murine isolate crosses rodent skin better than a human one and vice-versa (Benex & Bayssade-Dufour 1978). No sig-nificant difference in the number of cercariae maturing into adult worms (infectivity) was revealed, however, the human isolate yielded slight higher worm recovery. This result is in agreement with earlier laboratory investigations that have shown analogous data using C3H mice (Bastos et al. 1979). Although no statistically significant, sex-ratio was male biased, as previously reported in either naturally infected N. squamipes trapped in Sumidouro (Silva et al. 1992) or C3H mice infected with isolates derived from Vale do Rio Paraíba do Sul, São Paulo, Brasil) (Bastos et al. 1979). The normal adult worm sex-ratio in Brazil is male biased (Souza et al. 1996). The origin of this bias is not well understood. Under controlled laboratory conditions, it was clearly demonstrated that male and female cercariae show differences during life-cycle: while females had a higher performance regarding production, longevity and snail size, male cercariae are more infective to vertebrate hosts. In contrast, experimental mixed infections indicate that cumulative male and female cercariae infectivity is greatest when the sex-ratio is balanced. It is suggested that male presence stimulates female infectivity (Boisser & Moné 2000). Recent studies have shown a positive linear relationship between worm burden and male proportion (Boisser & Moné 2001). Results from the present work can't contribute in this respect because sex worm ratio is not statistically different. Evidence has been presented that adult worms have a realized niche (portion of the fundamental niche occupied within the host) for egg-laying (Holmes 1990). In this regard, about 50% of the eggs lodge in the distal segment of the small intestine (Costa & Katz 1982). Nevertheless, the pattern of egg distribution in the large intestine was slightly unequal between SR and SH isolates. These later findings may be the result of a difference in the wandering capacity inside the portal system vessels between both sub-populations (Valadares et al. 1981). Adult male and female schistosomes reside paired so that the female lies in a ventral groove (gynaecophoric canal) of the male (Skelly et al. 1998). It could point out that the female transportation toward the egg-laying site is subordinated to males' complex and well-developed musculature (Mair et al. 1998, Morand & Muller-Graf 2000). Inquiringly, a foregoing biometric study regarding the same isolates herein studied have evidenced that the rodent isolate has larger adult male worms than the human isolate (Machado-Silva et al. 1994). One could speculate that small size impairs male migration and therefore, reduction in number of trapped eggs. However, this was not herein detected. In this study, the isolates presented only significant biological differences regard parasite-snail relationship. Given that digenetic trematodes are primitively snail parasites and not of vertebrates perhaps their role as a biological filter is more efficient than vertebrate hosts (Freire et al. 2002). Phenotype and genotype studies have suggested that S. mansoni sympatric populations (murine and human) belong to a same population, however, genetic diversity within and among individual hosts have been recently described (Sire et al. 2001). Of particular relevance is the fact that both isolates share phenotypic characteristics (Machado-Silva et al. 1994, Neves et al. 1998, Freire et al. 2002). This observation is coherent with the results obtained with sympatric isolates from Guadeloupe and Senegal when a biochemical marker (isoenzymes profile) was used (Sène et al. 1997). Molecular studies have revealed that Brazilian allopatric strains (rodent and human) share the same mitochondrial DNA type (Desprès et al. 1993). More recently, a genetic similarity between strains isolated from humans and murines in Africa was also recorded (Duplantier & Sène 2000). It is perhaps relevant to mention that our data provide support to the hypothesis that in a same biotope, mixed populations or sub-populations circulate among their main host (human or rodent) that act as a biological filter (Freire et al. 2002). As genetic differentiation within infra-populations in rodents has been found (Sire et al. 2001), DNA microsatellite markers could be used to investigate the genetic structuring of natural populations (Durand et al. 2000, Rodrigues et al. 2002). Finally, the results presented here support the need for further investigations into the possible selective pressures acting on both populations. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc03159t1.jpg] |
| |||||||||

{kind=link}