 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
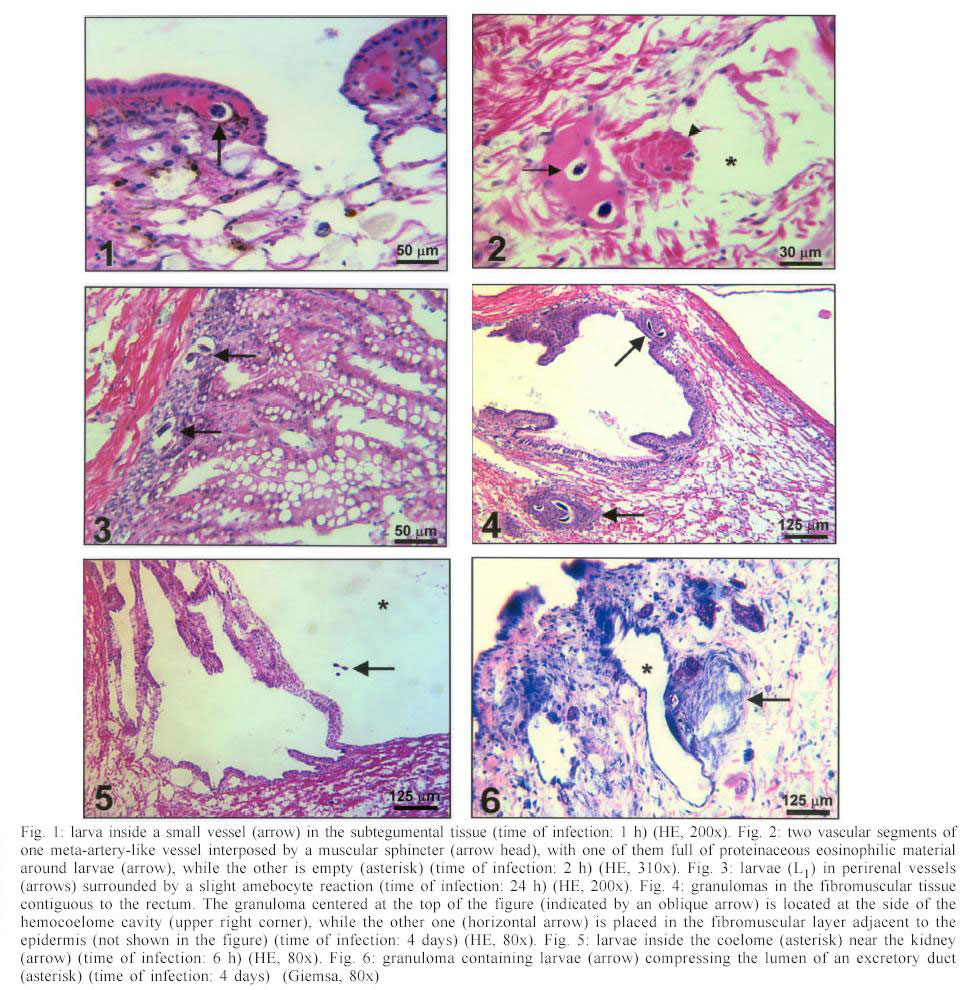
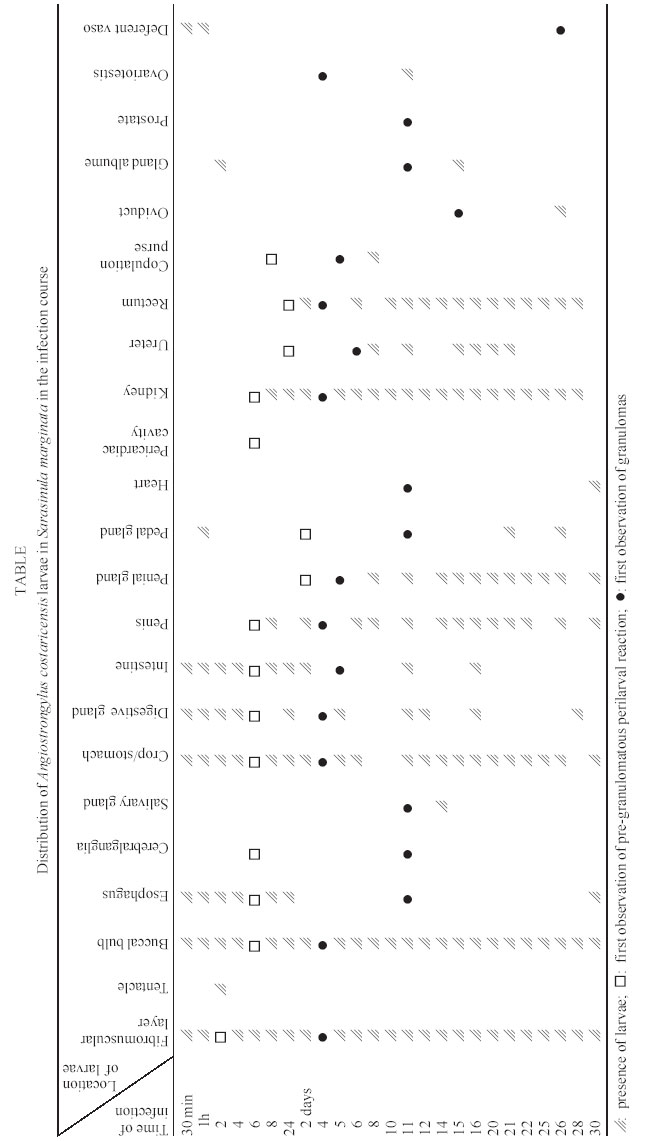
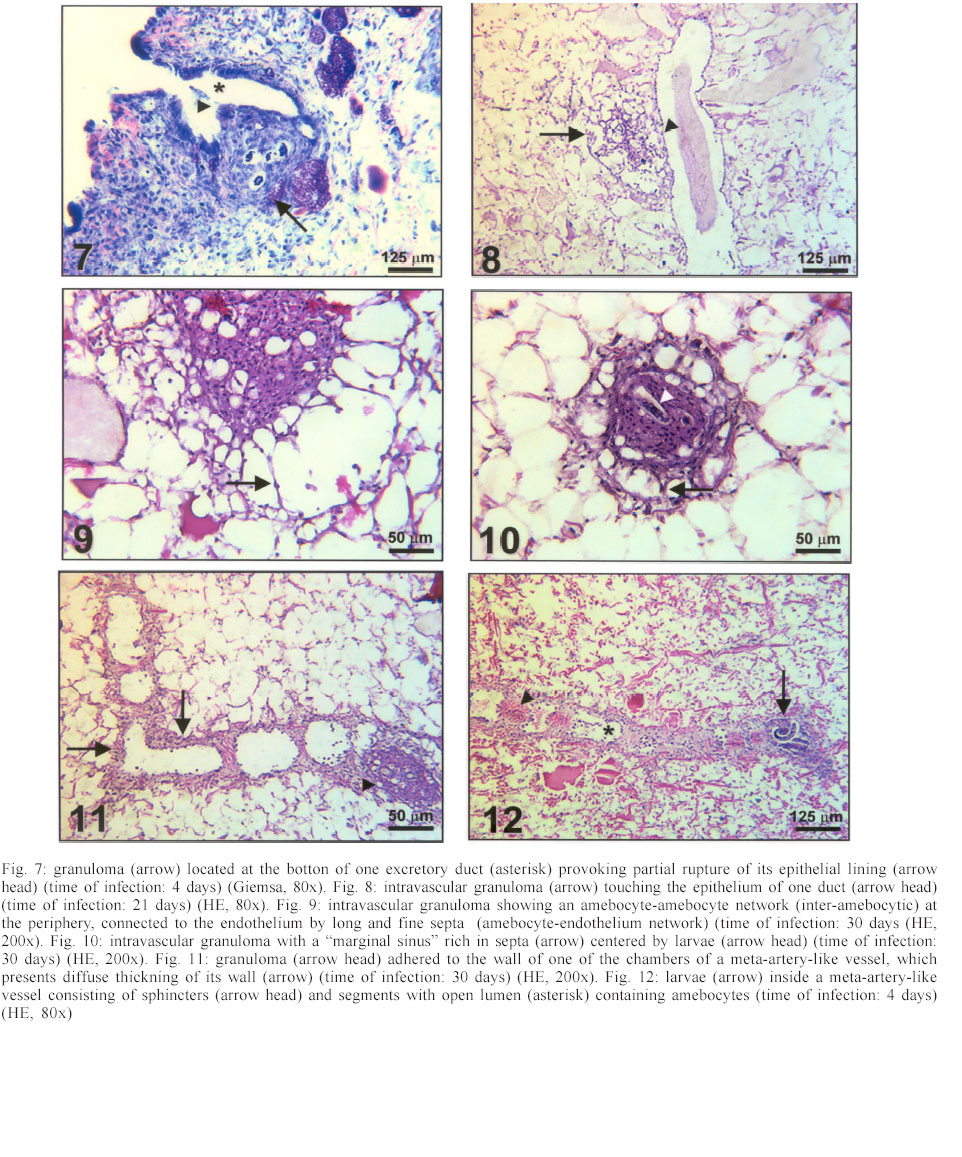
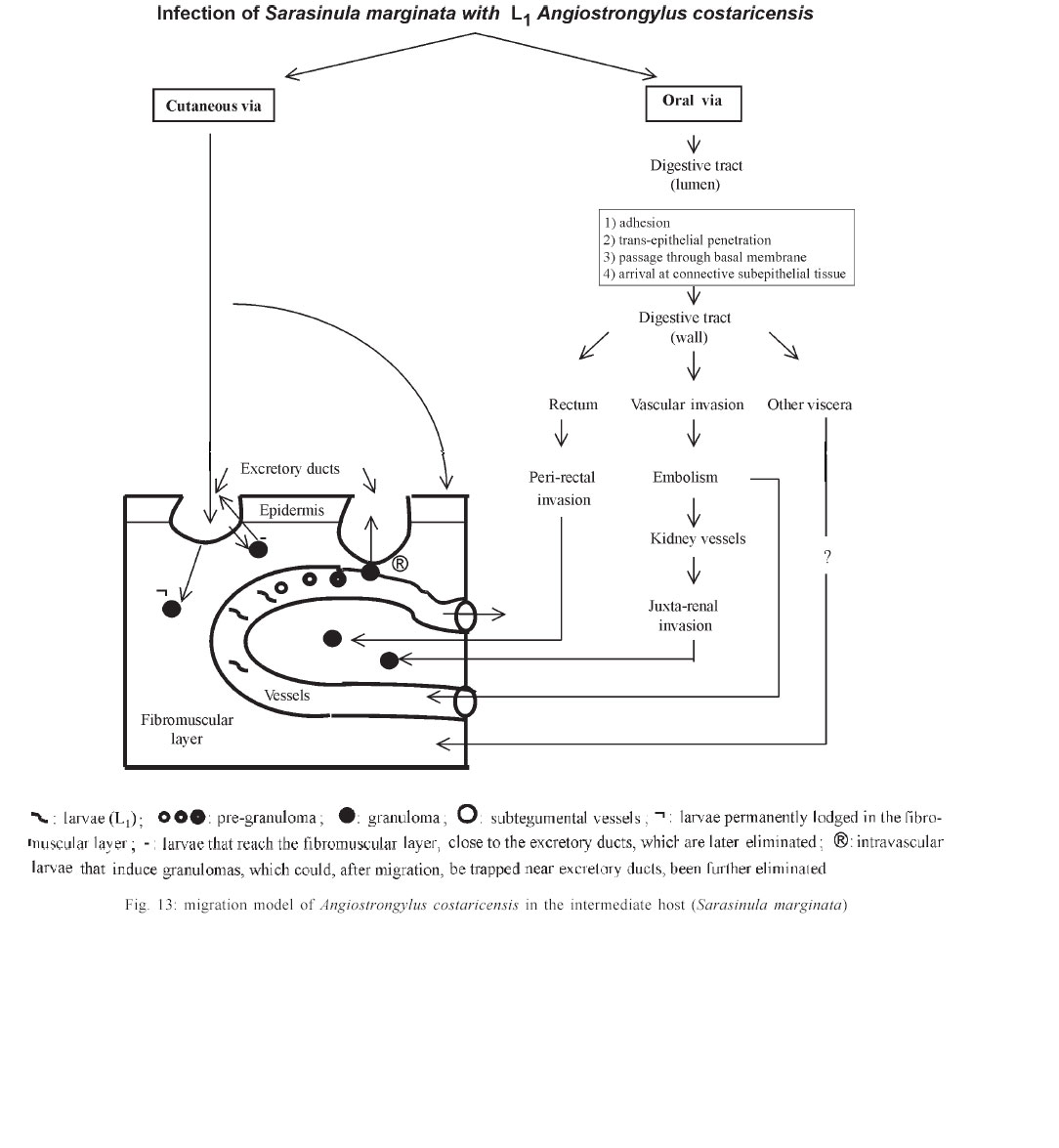
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 7, Sept, 2003, pp. 893-898 Angiostrongylus costaricensis and Experimental Infection of Sarasinula marginata II. Elimination Routes Cristiane Lafeta Gomes Furtado Mendonça, Omar Santos Carvalho, Ester Maria Mota*, Marcelo Pelajo-Machado*, Luzia Fátima Gonçalves Caputo*, Henrique Leonel Lenzi*/+ Centro de Pesquisas René Rachou-Fiocruz, Belo Horizonte, MG, Brasil *Departamento de Patologia, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil Partial financial support: Papes 383-Fiocruz Received 17 September 2002 Code Number: oc03177 Angiostrongylus costaricensis intermediate hosts are terrestrial mollusks mostly belonging to the Veronicellidae family. In the present investigation we focused on the mechanisms of larval expulsion from Sarasinula marginata infected with A. costaricensis. Twenty-five mollusks were individually infected with 5000 L1 and sacrificed at 30 min and 1, 2, 4, 6, and 8 h post-infection and at days 1, 2, 4, 5, 6, 8, 10, 11, 12, 14, 15, 16, 20, 21, 22, 25, 26, 28, and 30 post-infection; the mollusks were then fixed and stained. Diverse organs involved throughout the course of the migratory routes of larvae from oral penetration on were specified and the mechanisms of larval access to the fibromuscular layer through the kidney, rectum, and vascular system were defined. The elimination of L3, derived from oral and/or cutaneous infections, appears to depend on granulomas located close to the excretory ducts of mucous cells. Key words: Angiostrongylus costaricensis - Sarasinula marginata - Veronicellidae - migratory routes - larval elimination - amebocyte - invertebrate granuloma Angiostrongylus costaricensis (Morera & Céspedes 1971a) is the aetiological agent of abdominal angi-ostrongyliasis. Its life cycle in invertebrate hosts begins with ingestion of the larval first stage (L1) by the mollusk and/or percutaneous penetration by the larva, followed by two moultings (L1/L2/L3) in the fibromuscular layer and, exceptionally, inside the viscera. Larvae L3 are eliminated together with the mucous secretion of the mollusk (Morera & Céspedes 1971b, Conejo & Morera 1988, Thiengo 1996). In our previous work, studying the life cycle of A. costaricensis in Sarasinula marginata (Semper, 1888), we confirmed the simultaneous occurrence of oral and percutaneous infection of mollusks and described in detail the mode and sites of L1 penetration into the mollusk. The involvement of several organs during the migration of the parasite in the host tissues was also reported and the cellular perilarval (pre-granuloma and granuloma) and systemic (hemocytosis) reactions in the mollusk were characterized (Mendonça et al. 1999). It is important to stress that this mollusk has never been found naturally infected. Here we focus on the probable mechanism, which promotes larval elimination together with the mollusk's mucous secretion, and we also add more details to the previous report better clarifying the migratory routes of A. costaricensis in the intermediate host. MATERIALS AND METHODS Twenty-five S. marginata slugs captured in vegetable gardens in the city of Rio de Janeiro, where no evidence of natural infection was detected, weighing about 1 g, were individually kept in plastic containers and fasted for 5 days. They were then exposed to larvae L1 of A. costaricensis (Criciumal strain) by feeding on circular lettuce fragments (1 cm in diameter) covered with approximately 5000 larvae concentrated in 100 µl of a liquid solution of healthy mouse feces. The slugs were killed at 30 min, 1, 2, 4, 6, 8 h, and at 1, 2, 4, 5, 6, 8, 10, 11, 12, 14, 15, 16, 20, 21, 22, 25, 26, 28, 30 days after infection, and fixed in Carson's Formalin-Millonig (Carson et al. 1973). Serial cross-sections from mollusk's whole body were stained with Haematoxylin-Eosin (HE) and/or Lennert's Giemsa (Lennert 1978) and all slides (about 500 per slug) were examined by brightfield microscopy. For docu-mentation, selected slides were analyzed with a Zeiss photomicroscope III and the images were captured with a chilled Hamamatsu color 3 CCD camera (model C-5810), stored in tagged-image file format (TIFF) and printed with a Codonics TM NP-1600 photographic dye-sublimation printer or Deskjet 930C (Hewlett Packard). RESULTS After 1 h, L1 were occasionally detected inside peripheral vessels of the subtegumental vascular network (Fig. 1) and 2 h after infection they were also found into the vessels with intermittent muscular sphincters, located in the fibromuscular layer (Fig. 2), and in some narrow vessels at the renal periphery. After 4 h of infection, L1 reduced in number within the digestive tract lumen, being also found in the fibromuscular layer, buccal bulb, esophagus, crop, stomach, digestive gland, and intestine. From 6 h of infection on, larvae were mainly located in the fibromuscular layer and were also found in large numbers in the peripheral and interstitial portions of the kidney (Fig. 3) and rectum (Fig. 4). Few of them were detected in the same organs as described above in addition to the ureters and vessels eventually in the sheath of the penis, ovariotestis, oviduct, copulation purse, coeloma (Fig. 5), and salivary, pedal and penial glands. Almost all larvae were surrounded by a pre-granulomatous cellular reaction (Table). After the fourth day of infection, larvae (L1 and L2) were surrounded by typical granulomatous reactions. Some granulomas were compressing the ductal epithelium of mucous cells in the fibromuscular layer (Fig. 6) leading to its atrophy and local rupture (Fig. 7). On the fifth day, well-organized granulomas were also detected within vessels of the fibromuscular layer. This aspect pre-dominated from the 21th day of infection on. Some of the granulomas were positioned near the excretory ducts of the mucous cells (Figs 6, 7, 8). After 26 days of infection, the granulomas were mainly located intravascularly, adhering strongly to the endothelium through very thin septa (Figs 9, 10) creating a sort of "peripheral or marginal sinus" between their core and the endothelial covering. Partial obstruction of the vascular lumen by the granulomas caused a diffuse thickness of vascular walls (Fig. 11) and dilation of meta-arterial-like vessels (Fig. 12). These vessels were characterized by the intermittent alternation of muscular sphincters and segments with an expanded lumen (manuscript in preparation). In Fig. 13, we propose a general and simplified scheme based on the present and previous results about penetration sites (Mendonça et al. 1999) and migration routes of A. costaricensis in S. marginata, considering also the possible mechanisms of larval release to the external environment. DISCUSSION The current work detailed the migratory routes of A. costaricensis in S. marginata and the putative mechanisms of L3 expulsion from the mollusk's body. The involvement of different organs in the course of the migratory routes of larvae from oral penetration on was specified, pointing out the possible mechanisms of larval access to the fibromuscular layer through kidney, rectum, and vascular emboli. Together with Conejo and Morera (1988), we share the same hypothesis of L3 elimination as an accidental process in which random location near excretory ducts enables larval elimination through mucous secretion after mechanical rupture of the granulomas or perilarval amebocyte reactions. This fact might explain the low recovery of larvae from the mollusk's body, both in natural (Rambo et al. 1997) and experimental infection (Bonetti & Graeff-Teixeira 1998) since elimination is apparently independent of larval maturation and possibly depends on mechanic and accidental processes. It is relevant to remember that L3 are inactive, or immobile inside the mollusks, unable to achieve active elimination. Indeed, we have observed that some granulomas containing L3 were transported or formed close to excretory ducts, compressing their epithelium (Fig. 6) and occasionally provoking partial epithelial disruption (Figs 7, 8), thus facilitating L3 elimination. Lie and Heyneman (1976) similarly suggested that parasite encapsulation in the mollusk subepithelial fibromuscular layer resulted in occasional sporocyst elimination in the Echinostoma lindoenseBiomphalaria glabrata model. As amebocyte capsules became thicker, the strata close to the mollusk epithelium became thinner and turgid so that rupture of the epithelium allowed encapsulated sporocyst evasion through it. Richards and Merrit (1967) observed that such capsules persisted for long periods of time and viable Angiostrongylus cantonensis L3 were recovered from Biomphalaria up to 12 months post-infection. The larvae possibly acquired by cutaneous infection can hypothetically follow three different directions: (1) be kept arrested by the granulomatous reaction in the fibromuscular layer; (2) be lodged close to ducts, and (3) invade vessels, as is the case for larvae acquired by oral infection, being further transported intravascularly and trapped by granulomatous reactions adjacent to excretory ducts, from where they can be possibly eliminated. The current work also shows that the rectum was highly infected by the parasites. Except for the kidney (Mendonça et al. 1999) and rectum, few larvae were found within other viscera (Table). These two organs seem to be the preferential route for larvae that have orally penetrated to finally reach the fibromuscular layer. They were more intensely parasitized when compared to other organs in all slugs. The access of larvae to the fibromuscular layer was facilitated by the anatomical insertion of the kidney and rectum within it. In fact, larvae surrounding the rectum were found both in the coeloma and in the peripheral portion of the fibromuscular layer (Fig. 4). On the other hand, larvae, which penetrated the fibromuscular layer through the kidney, appeared to have been trapped as emboli in peripheral renal vessels, which are inside this layer ( Fig. 3). These results agree with observations made by Richards and Merrit (1967) and Harris and Cheng (1975) in the A. cantonensisB. glabrata model, where the importance of kidney and rectum as the main migration routes to the fibromuscular layer was emphasized. However, it was not possible to identify the final habitat of the larvae that reached the fibromuscular layer through those two mentioned organs; it is likely that they are retained in their surroundings. There is a remote possibility that the larvae, after leaving kidney and rectum, could also penetrate the vessels and/or migrate out of the vessels to the epidermal surface for further elimination. Frequent findings of larvae within the vessels (Figs 1, 2, 3) are indicative that larvae circulate until they are arrested inside the fibromuscular layer vessels by amebocytic reactions (Figs 8, 9, 10, 11, 12). According to these data, it is possible that larvae are arrested by the cellular reaction in the viscera (ectopic localization), not being eliminated through mollusk mucus. These larvae are epidemiologically important for life cycle maintenance in nature since they continue to develop even after being arrested and their elimination occurs after digestion in the vertebrate host's stomach. The location of larvae inside the vessels indicates that Angiostrongylid larvae prefer the intravascular habitat in the invertebrate host (Mendonça et al. 1999), or that they only form a passive embolus, since some larvae were detected in the heart and pericardiac cavity (Fig. 5). Most of the granulomas were well fixed to the endothelium by thin cellular septa formed by fibroblast-like cells from the amebocytic reaction (Fig. 10), leading to the formation of a "marginal sinus" causing thickening of vascular walls (Fig. 11) and dilatation of meta-arterial-like vessels (with segmented sphincters) (Fig. 12). Our study allows us to conclude that larval access to the fibromuscular layer via the oral route occurs by two different manners: (a) through kidney and rectum, and (b) by vascular emboli. Only those larvae that reached the fibromuscular layer through emboli, contributed to the parasite cycle, eliminating L3. Such elimination might depend on intravascular granulomas near the ducts. Larvae that prenetrate through the skin also seem to depend on granulomas located close to the excretory ducts of the mucous cells to be eliminated. ACKNOWLEDGEMENTS To AL de Amorim, FF Cruz, and ID Pedro (Department of Pathology, Instituto Oswaldo Cruz) for technical assistance. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc03177f1-6.jpg] [oc03177t1.jpg] [oc03177f7-12.jpg] [oc03177f13.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}