 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
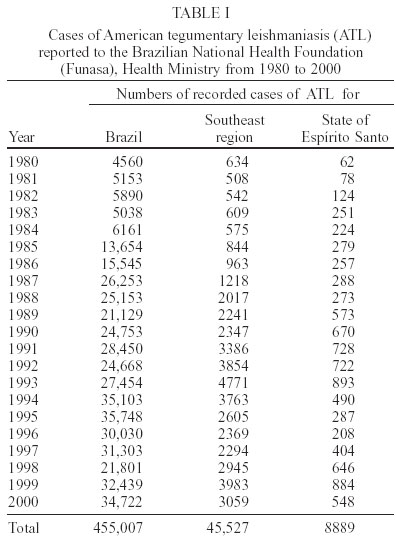
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 8, Nov, 2003, pp. 1003-1010 Epidemiological and Clinical Features of Leishmania (Viannia) braziliensis American Cutaneous and Mucocutaneous Leishmaniasis in the State of Espírito Santo, Brazil A Falqueto, PA Sessa, AL Ferreira, VP Vieira, CB Santos, JBM Varejão, E Cupolillo*, R Porrozzi*/**, LE Carvalho-Paes*, G Grimaldi Jr*/+ Unidade de Medicina Tropical, Departamento
de Patologia, Centro Biomédico, UFES, Vitória, ES, Brasil *Departamento
de Imunologia **Departamento de Ultraestrutura e Biologia Celular, Instituto
Oswaldo Cruz-Fiocruz, Av. Brasil 4365, Financial support: Pronex 3/CNPq/MCT (Brazil) Code Number: oc03196 Received 10 June
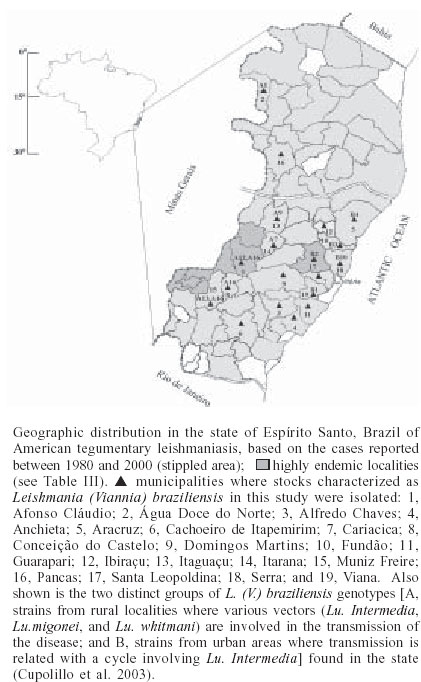
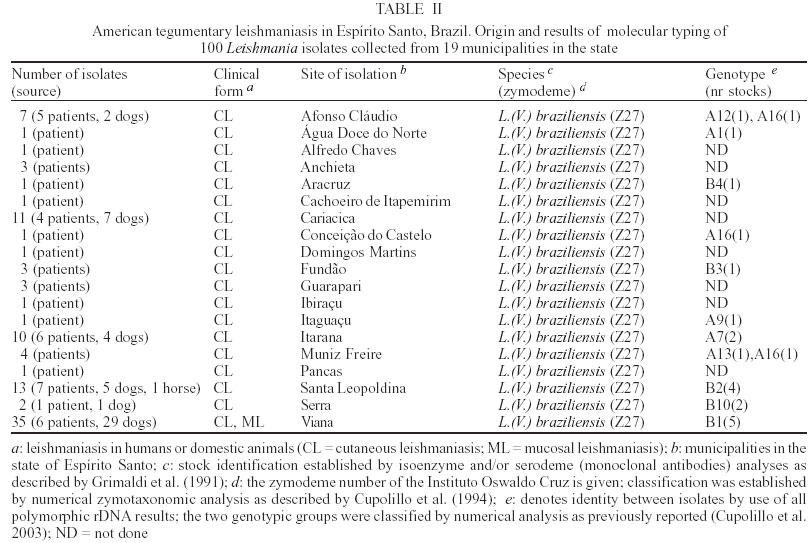
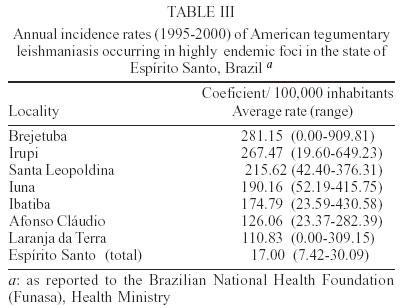
2003 Code Number: oc03196 Between 1985 and 2000, epidemiological surveys of the American tegumentary leishmaniasis (ATL) were carried out in several rural and urban communities in Espírito Santo, Brazil. A total of 100 stocks of Leishmania (comprising isolates from both human and canine hosts with ATL) were identified by two methods of molecular characterization, using specific monoclonal antibodies and multilocus enzyme electrophoresis. Parasite isolates from 19 municipalities were found to belong to the same zymodeme and serodeme type as of the Leishmania (Viannia) braziliensis reference strain. In contrast, our genotyping studies have shown intra-specific variation among these parasites (comparisons of the variability of the internal transcribed spacers between the small and large subunits of the rRNA genes of the 22 stocks studied revealed at least 11 genotypes). Two main clusters of L. (V.) braziliensis genotypes were observed, representing parasites collected from different endemic regions in the state, where transmission reflects distinct eco-epidemiological features. Infection with this pathogen was associated with the characteristic disease forms, but neither the clinical outcome nor the response to treatment could be related to the genetic polymorphism of the isolates, as defined by using the proposed methodology. Key words: Leishmania (Viannia) braziliensis - cutaneous leishmaniasis - molecular epidemiology - Espírito Santo - Brazil Leishmaniasis is a serious health problem in Brazil, where endemic regions have been spreading further a field and there has been a consistent rise in the numbers of recorded cases of American tegumentary leishmaniasis (ATL) in recent years (Table I). Control of leishmanisis in the New World is complicated by the variety of different Leishmania species (reviewed by Grimaldi & Tesh 1993), and by the fact that each of these parasites shows a unique epidemiological pattern (Lainson & Shaw 1987, Shaw 2002). Until effective vaccines are available, environmental-oriented control measures (such as vector and reservoir control and epidemiological surveillance) and chemotherapy of leishmaniasis will continue to be the best options for prevention and containment of the disease. Current problems however are (i) the fact that, although insecticides are quite useful in controlling phlebotomine sand flies in the domestic environment, sustained vector control utilizing large scale insecticide spraying in developing countries is costly and not feasible (Dye 1996); (ii) Leishmania spp. readily acquires resistance to antimonial drugs (Jackson et al. 1990, Grogl et al. 1992); and (iii) response to treatment varies considerable, probably depending on the parasite species involved and the clinical form or stage of the disease (Marsden 1985, Netto et al. 1990, Navin et al. 1992). From an epidemiological point of view, the various human pathogenic leishmanial species need to be identified and their geographic distribution and transmission cycle determined. In many endemic regions in the Americas, two or more parasite species are often sympatric (Grimaldi et al. 1989). Studies have also revealed genetic polymorphism in natural populations of different Leishmania species (Cupolillo et al. 1998, Mauricio et al. 2001, Ishikawa et al. 2002), which could explain the plasticity of these parasites and of their ability to adapt to changing ecological conditions (Cupolillo et al. 2003). Likewise, the intra-species variability could have a relationship with clinical pleomorphism of the disease (Guerbouj et al. 2001). In fact, many of these parasites are capable of producing in the human host a variety of clinical manifestations rather than a localized skin disease. For instance, a proportion of clinical infections leading to nasal metastasis is most associated with L. (Viannia) braziliensis (Grimaldi et al. 1987), but cases of this mutilating mucosal granulomata have also been reported as being associated with L. (V.) panamensis (Saraiva et al. 1985), L. (V.) guyanensis (San-trich et al. 1990), and L. (L.) amazonensis (Barral et al. 1991). Consequently, parasite identification is important for the diagnosis of the disease, and for the evolution of therapy and prognosis. ATL is endemic and widely distributed in the state of Espírito Santo, Brazil. The rapid increase in cases (Table I) coupled with evidence of transmission in the perido-miciliary environment (Ferreira et al. 2001) has prompted calls for adding preventive measures to the current national control policy of diagnosis and treatment. During the first epidemic, several cases of the disease were described from patients working in deforested areas or living in newly established communities (Machado 1913). However, environmental changes often modified the occurrence of ATL in the state, where new epidemics in non-forested areas were reported (Barros et al. 1985, Mayrink et al. 1985). Dogs and horses have been found infected with L. (V.) braziliensis and were implicated as domestic reservoirs of infection for humans in a number of foci (Falqueto et al. 1986, 1991). L. (L.) chagasi associated with visceral leishmaniasis (VL) in humans and dogs has also been recorded in the state (Grimaldi et al. 1989). Natural infections of spiny-rats (Proechimys iheringi) were detected in a secondary forest located in the municipality of Viana, but the three isolates were identified as a new enzymatic variant of L. (L.) forattinii (Falqueto et al. 1998), probably transmitted by Lutzomyia gasparviannai (Fal-queto et al. 1985). This report is an attempt to summarize in brief the data accumulated to date on the epidemiological and clinical aspects of the L. (V.) braziliensis ATL in Espírito Santo, including its known geographic distribution, zoonotic reservoirs, and insect vectors. Included are results of our present work (based on molecular typing of 100 parasite strains collected over a 15-year period from 19 municipalities in the state) as well as data from other recently published studies. MATERIALS AND METHODS Ecological description of the study areas - The ecology of ATL in Espírito Santo has been described in detail in previous publications (Falqueto et al. 1986, 1991, Ferreira et al. 2001). The transmission cycle of L. (V.) braziliensis occurs in areas situated at medium and lower elevations (50-750 m above sea level). The endemic foci are commonplace in small-scattered settlements, near mountains areas heavily deforested, but showing few remnants of the original vegetation. These rural villages are made up of well-dispersed houses and the space between houses is occupied by coffee and banana plantations. ATL is also found in old established communities close to important cities, like Vitória and Guarapari, on the Atlantic Coast. In the study areas (Figure), most transmission of the disease was in the peridomicile of houses; the maintenance cycle seems to involve domestic animal reservoirs (canine and equine) and phlebotomine sand flies with peridomiciliary habitats. Examination of patients - Clinical and laboratory ex-aminations of the subjects coming from different regions in the state, during the period from 1985 to 2000 were performed at the Medical Parasitological Section of the Federal University of Espírito Santo. All necessary in-formation on patients was recorded on registration cards prepared by the authors. House visits were also made in the endemic areas to search for unreported leishmanial cases. All consenting subjects with suspicious ATL were examined at the health posts, where specimens for smear and culture were taken for further detailed examination. Diagnosis was based on: (i) clinical appearance of the lesion; (ii) the positive Montenegro skin test (MST), indicating previous exposure to leishmanial parasites; (iii) the direct demonstration of the parasite in the lesion through examination of stained smears; and/or (iv) by isolating the parasite from tissue fragments, using in vivo and in vitro systems as described (Falqueto et al. 1991). The MST, which measures the cutaneous delayed-type hypersensitivity (DTH) reaction to Leishmania-derived antigens, was performed as described by Sokal (1975). The antigen (leishmanin), provided by the Fiocruz, Biomanguinhos Unit (Rio de Janeiro, Brazil), consisted of pooled heat-killed cross-species promastigotes suspended in PBS with 0.5% phenol. An induration diameter of equal or more than 5 mm was considered positive. Furthermore, analyses of specific cell-mediated immunity as detected by measuring in vitro lymphocyte proliferation and gamma interferon (IFN-γ) production] and humoral immune responses (measurements of specific antibodies) were performed in 10 subjects with either active ATL or cutaneous scars (in those who one or two months earlier had recovered from infection). The methods followed for peripheral blood leukocyte (PBL) preparations and assessment of recall proliferative and IFN-γ responses were those as described (Turetz et al. 2002). Cell proliferation was expressed as stimulatory index (SI) and was considered positive if > 2.5; IFN-γ in super-natants of cultures of stimulated cells were measured by ELISA. Antigen-specific serum antibody concentrations were determined using a standard ELISA technique (Badaró et al. 1986). The soluble leishmanial antigens (SLA) from L. (V.) braziliensis promastigotes was made as described (Amaral et al. 1996), and used at different concentrations for ELISA and blast trans-formation assays. Isolation of parasites - Primary isolation was made by culture in vitro of samples from the cutaneous lesions of hamsters, previously inoculated with the material obtained by lesion biopsies from patients or domestic animals with ATL. Stocks were initially cultivated in NNN blood-agar medium overlaid with Schneider's Drosophila medium (Sigma), supplemented with 10% heat-inactivated foetal bovine serum, FBS (Grimaldi et al. 1989). Positive culture containing promastigotes were expanded for growth in Schneider's medium containing 20% FBS and incubated at 25oC. Parasite characterization - The World Health Or-ganization (WHO) recommended reference strains for Leishmania species and parasite isolates used in this study are maintained at the DIFIOCRUZ Leishmania Type Culture Collection (registration no. 731, at the WFCC World Data Center on Microorganisms Directory). Stocks were stored as stabilates at _190oC, in liquid nitrogen containers, using 8% glycerol in Schneider's medium containing 30% FBS. Monoclonal antibodies used for typing leishmanial parasites (serodeme analysis) were group- or species-specific for members of both Viannia and Leishmania subgenera and have been described before (Grimaldi et al. 1987, 1989, 1991). Characterization of the Leishmania was performed with an indirect radio-immune binding assay, using whole parasite lysates as antigen, as report previously (Grimaldi et al. 1987). All stocks used in this study were characterized by multi-locus enzyme electrophoresis (MLEE). The methods used to prepare samples and study the electrophoretic mobility of some enzymes in agarose gels were performed as previously reported (Cupolillo et al. 1994). Samples were tested for the activity of the following enzymes: aconitate hydratase (ACON, E.C.4.2.1.3), glucose-6-phosphate dehydrogenase (G6PDH, E.C.1.1.1.49), glucose phosphate isomerase (GPI, E.C.5.3.1.9), isocitrate dehydrogenase NAD and NADP (IDHNAD & IDHNADP, E.C.1.1.1.42), malate dehydrogenase (MDH, E.C.1.1.1.37), malic enzyme (ME, E.C.1.1.1.40), manose phosphate isomerase (MPI, E.C.5.3.1.8), nucleosidase (NH1 & NH2, E.C.3.2.2.1), 6-phospho-gluconate dehydrogenase (6PGDH, E.C.1.1.1.43), phospho-glucomutase (PGM, E.C.1.4.1.9), leu-pro dipeptidase (PEPD, 3.4.13.9), and leu-gly dipeptidase (PEP2, E.C.3.4.11.1). Leishmanial isolates with the same electromorphs were classified into the same zymodeme (Cupolillo et al. 2003). Experiments using genomic restric-tion fragment length polymorphism (RFLP) analysis of the internal transcribed spacer (ITS) between the small (SSU) and large (LSU) subunits of the rDNA locus (PCR-RFLP ITSrDNA) of selected parasite strains were done to examine genetic polymorphism in this group. High resolution electrophoresis using the Genephor® apparatus was carried out in 12% polyacrylamide gels following the manufacturer's instructions (Amersham Pharmacia Biotech). The restriction enzymes used were those found to produce clear distinct banding patterns useful for grouping the isolates in genotypes (each genotype grouped isolates with the same banding pattern for all the restriction enzyme used) and were subjected to numerical analysis, as described elsewhere (Cupolillo et al. 1995, 2003). RESULTS Parasite characterization - Table II shows results of parasite identification, correlated with some epidemiological and clinical data, of the 100 Leishmania stocks isolated, classified according to their origin and geographic distribution. For comparison, in the characterization of the parasites, the WHO recommended reference strains were also included. Based on the characteristic reactivity of specific monoclonal antibodies (Grimaldi et al. 1987, 1991), we were able to characterize and classify 62 of the 100 Leishmania isolates (31 from human cases and 31 from infected dogs) collected in various municipalities. The apparent antigenic similarity among isolates was demonstrated using a large panel of cross-reacting monoclonal antibodies. These strains presented reactive patterns very similar to that of the L. (V.) braziliensis reference strain (MHOM/BR/1966/M2903), therefore they were identified as this parasite species. Results from agarose gel electrophoresis confirmed the specific identification made with the serodeme analysis for all isolates. The strains also had the same enzymatic profile classified as zymodeme IOC/Z27, which included the reference strain. In contrast, comparisons of the PCR-RFLP ITSrRNA of the 22 stocks studied (Table II) revealed at least 11 genetically variant strains (genotypes), each representing parasite(s) with unique ITS fragment profiles. Most of the genotypes were seemingly unique and localized to a particular endemic focus, but genotype A16 contained some isolates that were made from both human and canine hosts in three study areas. It is also of interest that some L. (V.) braziliensis isolates from cases of cut-aneous (n = 4) or mucosal (n = 1) disease represented the same genotype (B1). Prevalence and geographical distribution - ATL represents an increasing public health threat in Espírito Santo, where new foci are continually emerging; during 1995-2000 there was an incidence rate of 7.4-30.1 per 100,000 inhabitants (it is estimated 460 new cases annual-ly in a population of 3 million). However, it is likely that the numbers of recorded cases of ATL for the state (Table I) is still underreported. Furthermore, frank disease represents only a proportion of total infections as detected in the municipality of Itarana (Falqueto et al. 1991). The known geographic distribution in the state of ATL associated with L. (V.) braziliensis (based on clinical descriptions of cases and parasite isolations) is summarized on the map shown in the Figure. The disease occurs up to 850 m elevation; foci have been found in nearly all municipalities. ATL is highly prevalent in the mid-west and southeast regions of the state; most of the cases were reported from Afonso Cláudio, Brejetuba, Ibatiba, Irupi, Iuna, Laranja da Terra, and Santa Leopoldina, where recent epidemics have been recorded (Table III). Mucosal disease is relatively common, occurring in up to 15% of the cases. L. (V.) braziliensis showing a homogenous pattern either by serodeme analysis with specific monoclonal antibodies or by zymodeme analysis were detected in 19 of the 78 municipalities. These data confirm preliminary results on the presence of this parasite in some endemic areas (Falqueto et al. 1986, 1991), but also extend its geographic distribution in Espírito Santo. In addition, both L. (V.) braziliensis (based in this report) and L. (L.) chagasi (Grimaldi et al. 1989) can occur simultaneously within common endemic foci, such as those located in the municipalities of Pancas and São Gabriel da Palha. In the distribution of the 11 L. (V.) braziliensis genotypes, most of them were specific to some geographic localities, but one genotype (A16) was distributed in three municipalities (Afonso Cláudio, Conceição do Castelo, and Muniz Freire) which are contiguous geographic locations. Moreover, at least two distinct genotypes of this parasite were localized to a particular study area, such as in Afonso Cláudio (A12 and A16) and Muniz Freire (A13 and A16). Clinical characteristics of patients - A total of 1217 persons with ATL was examined (Table IV). Their disease was acquired in many different endemic foci in the state. In this group of infections, a variety of L. (V.) braziliensis genotypes were represented (Table II). Although the infections affected people of both sexes and of all age groups, the maximum incidence of the cutaneous disease occurred in the second decade of life. The number of lesions per person ranged from 1 to 6 (except for 14 patients with multiple skin lesions), but single skin ulceration occurred in most (65%) of the 1116 ATL patients examined. The characteristic ulcerous lesions with raised margin produced by L. (V.) braziliensis were most common (91.1% of the cases) on the lower and upper limbs (62.7%), the head and neck were affected in 24.9% of the patients while 12.4% had lesions on the trunk. Infiltrative plaque-like (7.5%) and verrucose warty (1.4%) lesions were occasionally seen. The majority (70%) of patients with ML (n = 101) were in their 50's and 60's and had a cutaneous scar after clinical cure of a primary lesion; 12% had an active skin lesion associated with the mucosal damage at the time they were examined, but the remaining had no detectable signs of a previous ATL. Most of the patients had only one mucosal site affected (78.2%) and, in this case, generally located at the nose; the remaining presented multiple mucosal lesions (2, 3 or 4 sites affected). Nasal mucosa lesion was detected in almost all the cases, the other lesion sites being the palate, pharynx, and larynx. The larynx was involved in 3.9% of our patients, in general associated with multiple mucosal lesions. Clinically, the affected mucosae presented diffuse infiltration with an irregular and bulged surface. When the larynx was involved, lesions frequently affected the epiglottis, a process that results in a horse and weak voice. The study subjects recovered following antimonial therapy [who had received distinct courses of a regime of 17 mg of meglumine antimoniate (Glucantime, Rhodia Farma, Brazil) per kg body weight per day for 10 con-secutive days], but the response varied considerably depending on the clinical form or stage of the disease. Those cases with cutaneous lesions recovered most easily; the mean time to clinical healing in this group was 50.2 ± 17.3 days (versus 80 ± 23.9 in the group with ML). However, similar levels of relapse were observed in both groups during the follow-up period (Table IV). These patients were further treated with higher doses, finally achiving cure. Side effects (such as local pain at the site of injection, arthralgias, astenia, mialgias, nausea and vomiting, haematological abnormalities, and/or alterations in electrocardiogram) were noted in a high proportion (about 80%) of treated patients. In the present study, we have evaluated both T cell-mediated and humoral immune responses of 10 subjects (aged from 12 to 72 years) who had acquired ATL in localities (Cariacica, Brejetuba, and Viana) of endemic L. (V.) braziliensis infection. Positive parasite-specific recall lymphoproliferative (the mean ± SD of SI = 6.5 ± 5.3; range 6-17) and IFN-γ (mean ± SD of IFN-g levels = 126 ± 74 pg/ml; range 90-320) responses were observed in all patients with active or healing disease (at the end of therapy), while these responses measured at short term (2 months) after cure from ATL were not above those of the healthy control individuals. The MST identified all the patients with active or healed ATL, although the reaction sizes varied among individuals tested (diameter of induration elicited by leishmanial antigens ranging from 8 to 30 mm) and were apparently not related with lesion development. Also the Ag-specific IgG antibody responses detected by ELISA in these patients were variable (the mean ± SD of absorbance values = 0.38 ± 0.37, range 0.14-1.2 at 1:640 serum dilution), but significantly (P < 0.05) above the serum levels (0.07 ± 0.02) found in healthy control subjects. DISCUSSION There is increasing evidence that some of the human pathogenic New World Leishmania can be maintained in both sylvan and urban transmission cycles. If so, the two cycles involve different vertebrate hosts. In Espírito Santo, the armadillo (Euphractus sexcinctus) and the paca (Agouti paca) are the presumed reservoir hosts in the sylvatic cycle of L. (V.) braziliensis (Falqueto 1995), whereas dogs and possibly equines (Falqueto et al. 1986, 1991) are the major vertebrate hosts in the peridomestic cycle of this parasite. The most interesting evidence obtained in a survey carried out in the municipality of Itarana (Falqueto et al. 1991), on the role of dogs as a major reservoir of L. (V.) braziliensis infection for humans, came from the comparison between individuals with leishmanial infection and their cohabitation with dogs presenting cutaneous leishmaniasis. Furthermore, in-tervention measures (such as active case detection and prompt/effective treatment of patients; control of potential reservoir host by performing euthanasia of infected dogs; and vector control by insecticide spraying in domestic and peridomestic areas) undertaken in the same endemic focus reduced transmission/morbidity drastically, since no new case was detected during a 2 years follow-up. In this report, attempts to isolate parasites from cutaneous or mucosal lesions by the culture of biopsy specimens were successful in 51 human cases and 49 domestic reservoirs (48 Canis familiaris and 1 Equus caballus). All the isolates identified as L. (V.) braziliensis were detected in 19 municipalities, showing that this pathogen is responsible for human and canine cases of ATL in the state. There was a clear correlation between the human disease and the relatively high prevalence and wide distribution of the infection in dogs (48 isolates were made from infected dogs in six study sites). Perhaps the most interesting information resulting from the molecular characterization is the similarity of the stocks derived from both human and canine cases of ATL. The apparent homogeneity of these parasites was confirmed at the genotype level (taking in consideration that some isolates made from both human and canine hosts in three study areas represented the same genotype), suggesting a possible domestic reservoir of this parasite. L. (V.) braziliensis has been associated with a number of different sand fly species (reviewed by Grimaldi & Tesh 1993). It is unclear whether all of the incriminated sand flies are important vectors, or if some are merely incidental hosts. Incrimination of a particular sand fly species as a vector is difficult, since several criteria are required (Young & Lawyer 1987). Lu. intermedia is probably the main vector of L. (V.) braziliensis in Southeast Brazil, while Lu. migonei and Lu. whitmani are of secondary importance (Rangel et al. 1984, Mayo et al. 1998). A search for suspect vectors was conducted in the municipality of Afonso Cláudio, in the mid-west region of Espírito Santo (a sample of 13,363 specimens belonging to 28 species was collected). The five most abundant man-biting species collected were Lu. intermedia (24.3% of the total), Lu. migonei (22.3%), Lu. whitmani (15.4%), Lu. fischeri (14.9%), and Lu. monticola (5.8%). Analysis of the distribution of these species at different altitudes provided evidence that Lu. fischeri and Lu. monticola were not involved in the transmission of the disease, whereas Lu. migonei and Lu. whitmani might act as secondary vectors of L. (V.) braziliensis and Lu. intermedia was probably the principal vector of the parasite (Ferreira et al. 2001). Also in the municipality of Itarana, the results confirmed that Lu. intermedia is the most commonly found sand fly species inside the houses and in the peridomiciliary dependences (76%) and may be implicated in ATL transmission (Falqueto et al. 1991). Evidence from our genetic analysis (Cupolillo et al. 2003) indicated a geographic structuring for some L. (V.) braziliensis genotypes. The isolates from Espírito Santo (showing two separate groups of genotypes) were considered as being involved in two transmission cycles. One group of parasites comprising isolates from urban areas was less diverse, compared with the other group comprising isolates from rural areas. The clinical features of the ATL cases recorded here were similar to those reported in other studies (Marsden 1985, Saraiva et al. 1989). L. (V.) braziliensis caused a variety of clinical lesions and the overall prevalence of mucosal disease in the state appeared to be less than 10% of patients. Most of the patients had an ulcerated skin lesion, but there was variation in the size and depth of the ulceration and this influenced the response to treatment. Occasionally we observed satellite nodules around the main lesions that eventually evolved to the disseminated form of the disease (Turetz et al. 2002). The specific treatment certainly speeded healing, but response to antimonial therapy was variable, probably depending more on the immune status of the patient (Saraiva et al. 1989), rather then any particular parasite strain involved (Guerbouj et al. 2001). Our genotyping studies did not identify a genotype specifically associated with antimony refractoriness in those patients that did not responded well to treatment. The key question for L. (V.) braziliensis in this respect is to establish the minimum dose of pen-tavalent antimonial that will cure the skin lesion rapidly and prevent mucosal relapse (Marsden 1985). ATL caused by this pathogen in Rio de Janeiro responds well to antimony and, in either cutaneous or mucosal disease, high cure rates were obtained with 5 mg/kg/day of glucantime® (Oliveira-Neto et al. 2000). Conversely, we have also observed spontaneous healing in some patients who for various reasons have never taken specific treatment. Patients with active or healing disease so far studied exhibit anti-Leishmania antibodies and parasite-specific cell-mediated immune responses. Results similar to ours were previously reported, studying distinct leish-manial infections (Carvalho et al. 1985, Sassi et al. 1999). The association of L. (V.) braziliensis with mucosal lesions is interpreted as evidence of parasite determinants of disease expression. Our genotyping studies indicate that such a fine degree of biological diversity in Leishmania seems unlikely to be related with disease expression. A clonal population contained several isolates from humans that developed skin and/or mucosal lesions. Clinical data have also provided evidence for the participation of host factors (such as genetic background and immune response) in the outcome of leishmanial infection in humans. The magnitude of the immune responses and hypersensitivity to leishmanial antigens tend to be greater in individual with ML than those with cutaneous disease (Carvalho et al. 1985, Saraiva et al. 1989), but in this study comparisons between groups did not show significant difference for skin induration diameters (Table IV). In conclusion, the species definitely associated with ATL in Espírito Santo is L. (V.) braziliensis. The asso-ciation of certain genotypes of this pathogen with specific transmission cycles could be interpreted as evidence of an adaptation of different clones to the vector species involved in the study sites. The presence of the same genotypes in humans and dogs in the same area is suggestive that intra-specific transmission cycles (related with Lu. intermedia, Lu. migonei, and Lu. whitmani) could be defined using molecular technology. This information is expected to yield new approaches oriented towards elucidating transmission of the disease and eventually to improved parasite control. ACKNOWLEDMENTS To Dr Luiz Carlos Cecilioti da Cunha and the staff in the Fundacão Nacional de Saúde for collecting local records of notified ATL cases. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc03196t1.jpg] [oc03196t4.jpg] [oc03196f1.jpg] [oc03196t3.jpg] [oc03196t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}