 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 8,Nov, 2003, pp. 1039-1043 Molecular Differentiation of Angiostrongylus costaricensis, A. cantonensis, and A. vasorum by Polymerase Chain Reaction- Restriction Fragment Length Polymorphism Roberta L Caldeira, Omar S Carvalho/+, Cristiane LFG Mendonça, Carlos Graeff-Teixeira*, Márcia CF Silva*, Renata Ben*, Rafael Maurer*, Walter S Lima**, Henrique L Lenzi*** Laboratório
de Helmintoses Intestinais, Centro de Pesquisas René Rachou-Fiocruz,
Av. Augusto de Lima 1715, 30190-002 Partial financial support: Fapemig and Fiocruz Received 24 July
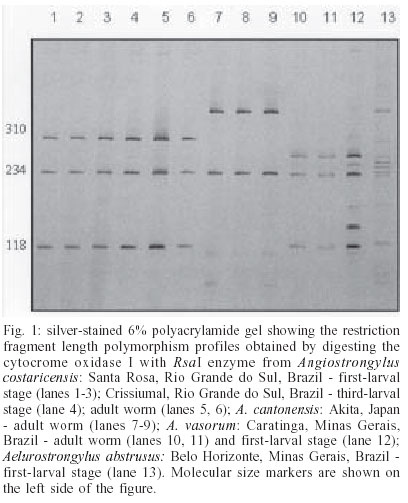
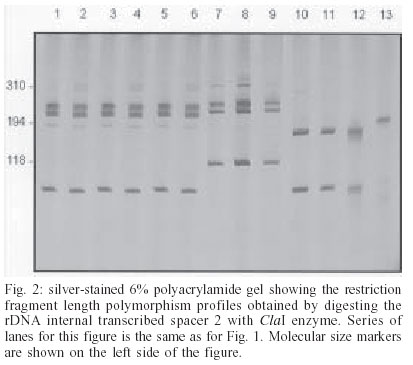
2003 Code Number: oc03203 Angiostrongylus cantonensis, A. costaricensis, and A. vasorum are etiologic agents of human parasitic diseases. Their identification, at present, is only possible by examining the adult worm after a 40-day period following infection of vertebrate hosts with the third-stage larvae. In order to obtain a diagnostic tool to differentiate larvae and adult worm from the three referred species, polymerase chain reaction-restriction fragment length polymorphism was carried out. The rDNA second internal transcribed spacer (ITS2) and mtDNA cytochrome oxidase I regions were amplified, followed by digestion of fragments with the restriction enzymes RsaI, HapII, AluI, HaeIII, DdeI and ClaI. The enzymes RsaI and ClaI exhibited the most discriminating profiles for the differentiation of the regions COI of mtDNA and ITS2 of rDNA respectively. The methodology using such regions proved to be efficient for the specific differentiation of the three species of Angiostrongylus under study. Key words: Angiostrongylus - polymerase chain reaction - restriction fragment length polymorphism - second internal transcribed spacer - cytochrome oxidase I Angiostrongylus (Nematoda: Angiostrongylidae) parasites may be found in the lungs and arteries of insectivores, rodents, canines, and felines (Anderson 1978). Some of these parasites may reach man as a definitive host, causing serious public health problems such as: eosinophilic meningitis caused by A. cantonensis (Alicata 1988), abdominal angiostrongyliasis by A. costaricensis (Moreira & Cespedes 1971), and diseases of the central nervous system by A. vasorum (Eckert & Lämmler 1972). Rodents are definitive hosts of A. cantonensis and A. costaricensis, whereas A. vasorum is a canine parasite. A wide variety of molluscs are intermediate hosts of such angiostrongylids that are frequently confused with other nematodes (Ash 1970). The occurrence of such parasites in humans emphasizes the importance of a precise differentiation among species. However, the specific identification of larvae and adult worms concerning Angiostrongylus genus based on morphological characters is unfeasible due to vague and similar descriptions on size and body shapes among species (Santos 1985, Ubelaker 1986). Indeed, correct morphological identification has been also precluded as an accurate diagnostic tool for many other organisms in which polymerase chain reaction together with restriction fragment length polymorphism (PCR-RFLP) have been used. This methodology provides an alternative approach for molecular differentiation and has been successfully employed to identify Biomphalaria snails (Caldeira et al. 2000, Vidigal et al. 2000), to distinct cryptic species within the Trypanosoma brucei group (Agbo et al. 2001) and also to separate closely related nematodes (Newton et al. 1998, Gasser et al. 1999, Wu et al. 1999, Otranto et al. 2001). In addition, sequencing of the second internal transcribed spacer region (ITS2), as well as PCR-RFLP has shown to be a powerful genetic tool for studying some Meta-strongylidae species (Leignel et al. 1997, Conole et al. 1999). The aim of the current study was to develop an appropriate methodology based on PCR-RFLP of the mitochondrial-derived cytochrome oxidase I gene (COI) and the genomic DNA based ITS2 of rDNA in order to differentiate A. costaricensis, A. cantonensis, and A. vasorum. These molecular markers were chosen based on their usefulness for discrimination of closely related species (Brown et al. 1979, Hillis & Dixon 1991). In addition, RFLP patterns from Aelurostrongylus abstrusus were compared with such species due to their taxonomic closeness with the genus Angiostrongylus. MATERIALS AND METHODS Parasites and DNA extraction - Adult worms as well as first and third-larval stages of A. costaricensis, A. cantonensis, and A. vasorum, kept under laboratory conditions, were used (Table). A. abstrusus was included in the study in order to provide restriction pattern data for further comparison with Angiostrongylus, due to their closeness. Adult worms were recovered from several sites of vertebrate hosts (lungs and mesenteric arteries); the first-larval stage was collected from feces of vertebrate hosts and the third-larval stage was obtained after pepsin digestion of the slug Sarasinula marginata. All parasites under study were sent to the Intestinal Helminthiasis Laboratory of René Rachou Research Center, Fiocruz (Brazil), in 70% ethanol solution, except for the specimens of A. cantonensis, which were sent lyophilized. Total DNA was extracted from worms and pools of larvae, using the phenol-chloroform method as described by Simpson et al. (1982). mtDNA COI amplification - A portion of the COI region was amplified using LCO (forward; 5'- ggtcaacaaatcataaagatattgg-3') and HCO (reverse; 5'-taaacttcagggtgaccaaaaaatca-3') as primers. These primers were designed upon evolutionarily conserved regions of the COI sequence (Folmer et al. 1994). PCR amplification was undertaken in a total volume of 10 µl consisting of: 1-10 ng of template DNA, 10 mM Tris-HCl, pH 8.5, 200 µM each dNTP, 1.5 mM MgCl2, 0.8 U of Taq DNA polymerase (Cenbiot, RS, Brazil), 50 mM KCl, together with 0.3 pmol of each primer. The reactions were covered with a drop of mineral oil and subjected to the following cycle program: initial de-naturation step for 5 min at 95°C, and then 40 cycles as follows: annealing at 50°C for 1 min, extension at 72°C for 2 min, denaturation at 95°C for 45 s and a final extension step at 72°C for 5 min. rDNA ITS2 amplification - The ITS2 region was amplified using the primers NC1 (forward; 5'-ACGTCTGGTTCAGGGTTGTT-3') and NC2 (reverse; 5'-TTAGTTTCTTTTCCTCCGCT-3') (Gasser et al. 1993) and PCR reaction was carried out as described above. The conditions in a thermocycler were: initial denaturation step for 90 s at 94°C, and then 39 cycles for: annealing at 58°C for 1 min, extension at 72°C for 90 s, denaturation at 94°C for 50 s and a final extension step at 72°C for 10 min. For both amplifications, a negative control (no template DNA) was included in all experiments. Three microliters of the amplified products were visualized on 6% silver stained polyacrylamide gels to have the quality of amplification checked. RFLP analysis -The PCR products were diluted in water, divided into 10 µl aliquots and then separately digested with different restriction endonucleases. Such endonucleases were randomly selected once COI and ITS2 sequences from Angiostrongylus are not available in the GeneBank. Six restriction enzymes were used: four base cutters RsaI, HapII, AluI, and HaeIII; the five base cutter DdeI and the six base cutter ClaI. Ten to 12 units of each enzyme were used for each digestion reaction, together with 1 µl of the respective enzyme buffer and 10 µl of the diluted amplification product. The digestion was performed for 3.5 h at 37°C and products were evaluated by visualization on 6% silver stained polyacrylamide gels, after phenol/chloroform extraction to remove protein. The results were recorded with a digital photodocumentation system (Mavica, Sony). A control of activity for each enzyme was performed by digesting 150 ng of pUC18 simultaneously with the samples under evaluation. RESULTS mtDNA COI - The amplification generated a fragment of approximately 700 bp. After enzyme digestion, all samples had their profiles reproduced and only some specimens of each species are demonstrated here. Out of the six enzymes, only HaeIII showed no restriction sites in the amplified fragment (data not shown). Through the other enzymes, it was possible to distinguish among the species under study (data not shown) and RsaI presented the best profile for differentiating worms and larvae (Fig. 1). rDNA ITS2 - PCR amplification yielded fragments ranging from 600 bp (A. costaricensis and A. vasorum) to 650 bp (A. cantonensis). After enzyme digestion all samples exhibited reproducible profile and only some specimens of each species are shown here. DdeI enzyme presented no restriction sites for the species under study (data not shown); HaeIII, AluI, and MnlI also showed no restriction sites for A. cantonensis and A. vasorum (data not shown). The profiles produced by RsaI allowed us to distinguish among the three species, however A. cos-taricensis and A. vasorum genetic profiles showed to be alike. ClaI enzyme exhibited the best profile for differentiating the worms and larvae (Fig. 2). The restriction patterns for some species were complex and the sum of sizes of the bands produced upon digestion was not equal to the size of the undigested PCR products. For instance, in the patterns produced by RsaI for A. vasorum (Fig. 1) the sum of the sizes of two bands (180 + 72 pb) was not equal to the estimated 600 pb. It might be due to overlapping of similar size fragments produced by the enzyme. DISCUSSION The present study, using molecular markers, allowed us to differentiate the larvae and adult worms from A. costaricensis, A. cantonensis, and A. vasorum, contributing to reduce time-consuming morphological identification, which usually takes around 40 days. Routinely, for parasite identification, third-stage larvae are isolated from naturally infected snails and further used for oral infection of vertebrate hosts. After adult worm maturation and migration to the final habitat, they are then recovered and morphologically analyzed. This usual procedure, besides being a long lasting process, requires the sacrifice of the vertebrate host to have the adult worms recovered. The importance of fast and accurate diagnostic tools for diseases caused by nematodes lies on knowledge enhancement on prevalence and geographical distribution, as such parasites have nearly all gastropods as their intermediate hosts. Humans are accidental hosts, who does not eliminate eggs or larvae in the feces, and serological diagnoses show low sensitivity and specificity (Graeff-Teixeira et al. 1997, Geiger et al. 2001). Unfortunately, there is no definitive diagnosis for humans and the infection rates may be underestimated. Efforts have been made in order to enlarge studies on nematodes of medical and veterinarian importance. Gasser and Newton (2000) reported genomic and genetic in-vestigations on bursate nematodes and remarked that although PCR-RFLP is widely used for several nematodes, it has been little utilized in studies on strongylidae. Most works on Nematoda have chosen rDNA regions because they are useful genetic markers for studies on diagnosis, systematics, molecular evolution, and many other aspects. Thus, the application of molecular markers is widely used to differentiate closely related species and in many cases only a target region is used, depending on the proposed aim. All regions of the nuclear and mitochondrial genomes of parasites accumulate mutations over time and some regions are more accessible to nucleotide alterations than others (Gasser 1999). Anderson (2001), studying Ascaris epidemiological aspects, showed how molecular data, derived from a single DNA region, might lead to ambiguous conclusions. Therefore, in order to support our evidence, two distinct DNA regions were chosen, which provided us with convergent and consistent results. Furthermore, no intraspecific variation was detected in the regions ITS2 and COI, except for the larvae pool of A. vasorum (Fig. 1, lane 12), which presented a slight polymorphism in the COI region. This proposal is supported by the fact that there is a substantial variation in intra and interpopulational mtDNA as remarked by Tarrant et al. (1992) and Blouin et al. (1998). The ITS2 region exhibited few restriction sites regarding the applied enzymes used here. Out of six, only RsaI and ClaI found restriction sites for all species under study. Such data are in accordance with those by Romstad et al. (1998) who reported little or no variation in ITS2 sequences for strongylidae nematodes. Hoste et al. (1995) have also reported no variation in ITS2 sequences in Trichostrongylus eggs and/or worms. However, in both studies, the differences observed among species clearly exceeded the level of intraspecific variation, which reinforce the use of ITS2 region sequences to differentiate closely related species. On the other hand, Leignel et al. (1997) and Conole et al. (1999) reported a significant level of inter and intraspecific variations in the ITS2 sequences for Metastrongylus sister group of Angiostrongylidae. The primers set NC1-NC2 is known to amplify ITS2 from a variety of nematodes but not from other nematode groups or vertebrate host DNA (Newton et al. 1997, Hung et al. 1999). Preliminary studies, in our laboratory, showed that specific PCR directed to the regions ITS2 and COI from rDNA and mtDNA, respectively, for the following snails S. marginata, S. linguaeformis, and Biomphalaria glabrata, produced no fragments that overlapped those from nematodes studied here. In the present study, PCR-RFLP showed to be a fast and accurate diagnostic tool to Angiostrongylus of me-dical and veterinarian importance. Both genetic regions, ITS2 and COI, yielded satisfactory results considering the proposed aim. The most appropriate restriction enzymes for ITS2 of rDNA and COI of mtDNA were ClaI and RsaI, respectively. As already stressed, PCR-RFLP methodology has the advantage of producing clear and distinct profiles for each species. ACKNOWLEDGMENTS To Dr Kentaro Yoshimura, Department of Parasitology, Medical School of University of Akita, Japan, for providing us with Angiostrongylus cantonensis; Joziana Barçante, Department of Parasitology of Federal University of Minas Gerais, Brazil, for maintaining the life cycle of A. vasorum; Ester Maria Mota, Department of Pathology, Oswaldo Cruz Institute-Fiocruz, Rio de Janeiro, Brazil, for being in charge of A. costaricensis life cycle. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc03203t1.jpg] [oc03203f1.jpg] [oc03203f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}