 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 8, Nov, 2003, pp. 1059-1065 Redescription of Nyssomyia intermedia (Lutz & Neiva, 1912) and Nyssomyia neivai (Pinto, 1926) (Diptera: Psychodidae) José Dilermando Andrade Filho/+, Eunice A Bianchi Galati*, Alda Lima Falcão Laboratório
de Leishmanioses, Centro de Pesquisas René Rachou-Fiocruz, Av. Augusto
de Lima 1715, 30190-002 Financial support: Fundação Nacional de Saúde and Fiocruz Received 27 May
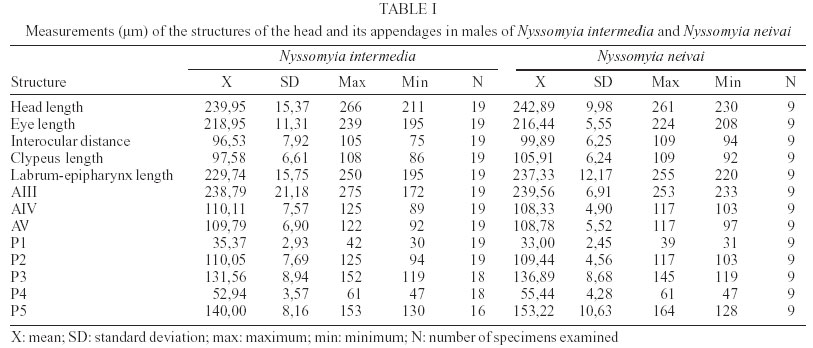
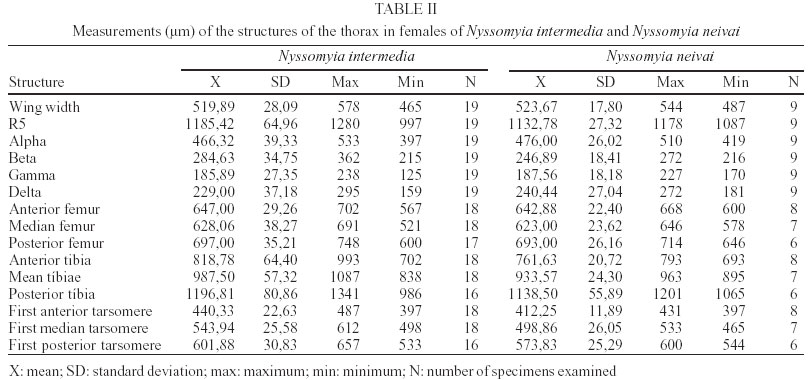
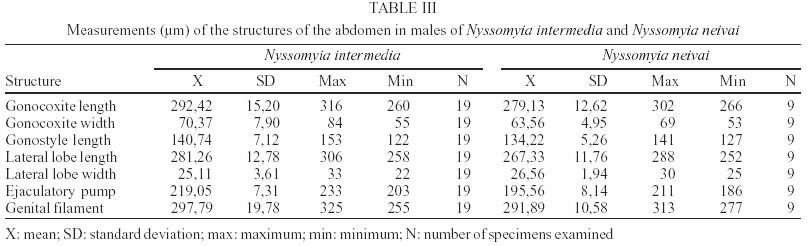
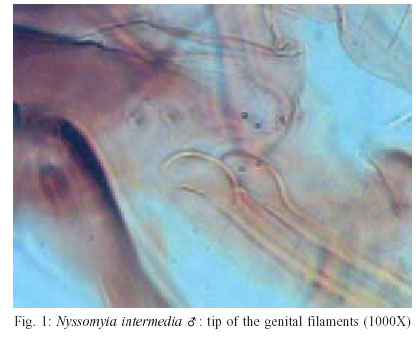
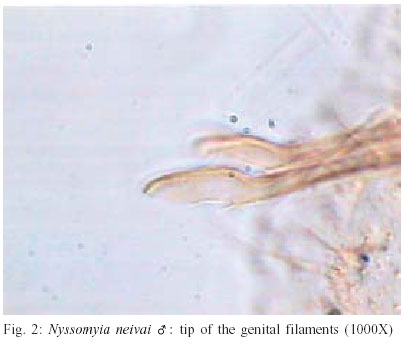
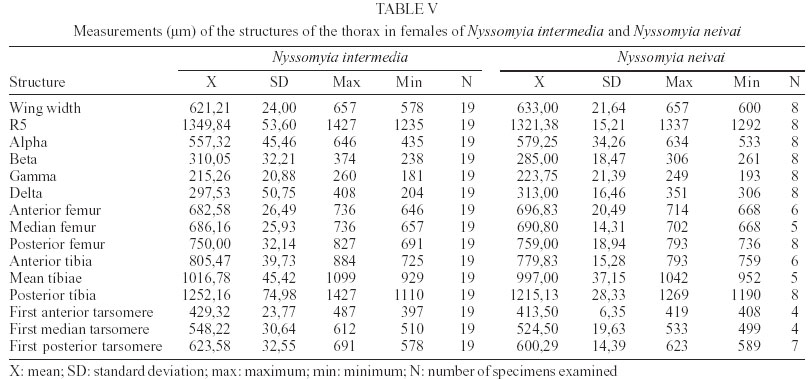
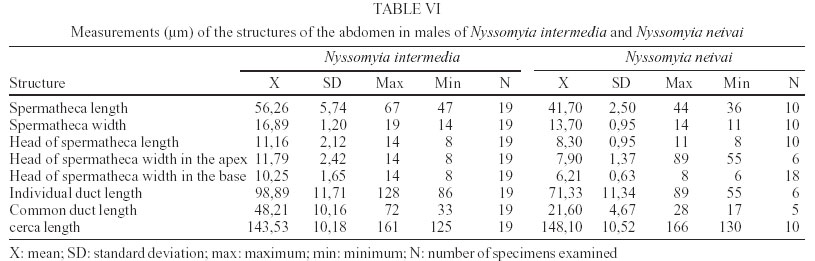
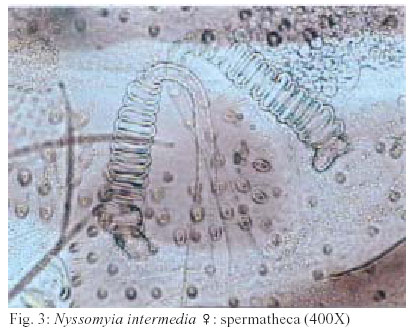
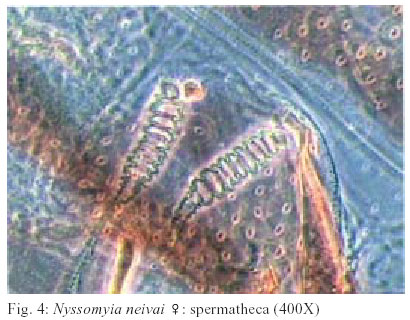
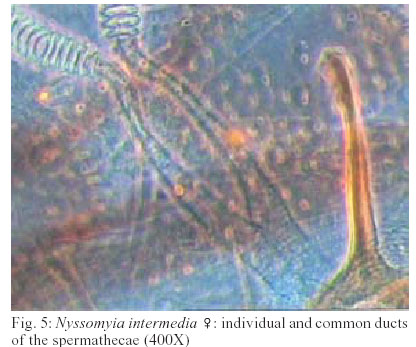
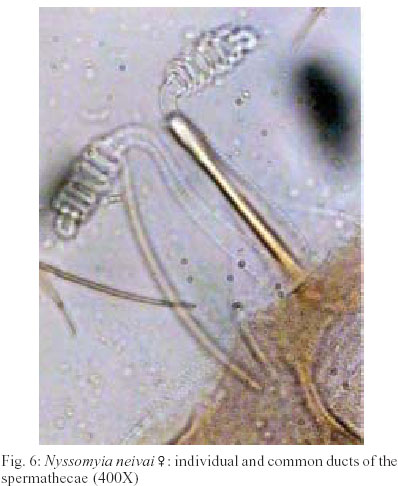
2003 Code Number: oc03207 The phlebotomine sand flies Nyssomyia intermedia and Nyssomyia neivai are the probable vectors of American tegumentary leishmaniasis in the Southern and Southeastern regions of Brazil. These species form a complex, being difficult to separate between either females or males of the two members based on recognized morphological characteristics. Both N. intermedia and N. neivai are redescribed here in the search for characters that facilitate their correct identification. It was possible to differentiate females by means of spermathecal characteristics. Males could be separated with confidence by the tips of the genital filaments, which have the form of a deep spoon, the angle of the concavity being well accentuated in N. intermedia and much shallower in N. neivai. Key words: Nyssomyia intermedia - Nyssomyia neivai - Phlebotominae - Psychodidae Nyssomyia intermedia was described from adults collected at Fazenda Ouro Fino, municipality of Além Paraíba, Minas Gerais. Material from the states of São Paulo and Rio de Janeiro was also examined (Lutz & Neiva 1912). In this description these authors mainly discuss characters obtained from examination of male specimens, mention of females being limited to the phrase "the single female and two male specimens were collected at Ouro Fino". Nevertheless Barretto (1947, 1961), Martins et al. (1978), Young and Duncan (1994) and Marcondes (1997) consider this to be the first formal description of the species. França (1920) and Marcondes (1996) redescribed N. intermedia, the former author from insects captured in southern Bahia and Vila Americana in the state of São Paulo, and the latter on those captured at the type locality. This description included designation of a neotype female. Pinto (1926) described Nyssomyia neivai from a male captured inside a house at the Instituto Butantan, municipality of São Paulo. This was later designated as a junior synonym of N. intermedia by Pinto (1930) a view held by most phlebotomine workers until Marcondes (1996) revalidated the name, based on the holotype and on a female collected at Fortim Campero in the department of Tarija, Bolivia. Distinction between the females of these two species is based principally by characters of the spermathecae (total length, number of annulations, form and length of head, and length of the common duct in relation to the individual duct) and head, i.e., number of horizontal teeth in the cibarium (Marcondes 1996). Measurement values for the wing venation (alpha length and alpha/beta ratio) of both sexes and the male ejaculatory pump/genital filament ratio were both significantly greater in N. neivai. Nevertheless, Marcondes and Borges (2000) were unable to distinguish between the males of these species with confidence based on their morphology and could only separate them using morphometric studies and analyse artificial neural network. MATERIALS AND METHODS The adult male and female structures of N. intermedia and N. neivai examined included some of those described by Galati (1995) based on recommendations of the CIPA Group (1991). The classification scheme used is that of Galati (1995). The specimens of N. intermedia examined were captured at its type locality, Além Paraíba (21o53'S and 42o42'W), while those of N. neivai included the male holotype described by Pinto (1926) and specimens reared in the laboratory to the F2 generation. These were descended from insects captured in Conchal (22o18'S and 47o13'W) in the Brazilian state of São Paulo and supplied by Dr Cláudio Casanova. All insects were mounted on glass slide using Berlese liquid. All the material was examined under an Olympus, model CH-2 optical microscope, with measurements made using an ocular micrometer, duly calibrated for this apparatus. The photographs were taken on a Zeiss microscope, and digitalised for the microcomputer using the KS300 program. The females of species were mounted in Berlese's media and the males in Canada balsam. RESULTS All measurements are given in micrometers, with the mean, standard deviation, and number of specimens examined in parentheses. Redescription of the males of N. intermedia and N. neivai N. intermedia Head (Table
I): ratio of lengths of clypeus/head 0.39 (0.41 ± 0.04; n = 19),
eyes/head 0.89 (0.91± 0.04; n = 19). Ratio Cervix: ventro-cervical sensilla absent. Thorax (Table II): notum and paratergite dark chestnut, pleurae clear. Bristles absent in anterior region of katepisternum. Proepimeral and superior anepisternal bristles present. Wings measuring 554 (520 ± 28; n = 19) wide, R5 1201 (1185 ± 65; n = 19) long. Principal wing indices: a (alpha) 532 (466 ± 39; n = 19); b (beta) 238 (285 ± 35; n = 19); g (gamma) 136 (186 ± 27; n = 19); d (delta) 283 (229 ± 37; n = 19). Tarsomeres III and IV with thin spines implanted at two levels, one apical and other median, these being more robust in some specimens. Femur of anterior, median and posterior legs measuring 657 (647 ± 29; n = 18), 634 (628 ± 38; n = 18), and 702 (697 ± 35; n =17) respectively. Tibia of anterior, median and posterior legs measuring 804 (818 ± 64; n = 18), 952 (988 ± 57; n = 18), and 1178 (1197 ± 81; n = 16) respectively. Tarsomere I of anterior, median and posterior legs measuring 441 (440 ± 23; n = 18), 521 (544 ± 26; n = 18), and 589 (602 ± 31; n = 16), respectively. Abdomen (Table III): absence of papillae on abdominal tergites. Gonocoxite: 297 (292 ± 15; n = 19) long by 75 (70 ± 8; n = 19) wide in its median part, without tuft of bristles in its basal region and with some elements distributed sparsely over the median region. Gonostyle: 147 (141 ± 7; n = 19) long, with four well developed spines, one being apical, one external superior implanted on the apical fourth, one external inferior and one internal. These two last implanted at the same level, a little further on than the middle of the structure. Pre-apical bristle absent. Paramere digitiform, wide at base and middle of the structure with a sharp point on the ventral region ventral. Groups of fine bristles present on the dorsal margin of the apical region and also the ventral margin, implanted in the widest region of the paramere. Lateral lobe length approximately equivalent to that of the gonocoxite, with ratio gonocoxite/lateral lobe 1.05 (1.04 ± 0.04; n = 19). Aedeagus conical. Ejaculatory pump with wide body. Genital filament short, ratio genital filament/ejaculatory pump 1.40 (1.34 ± 0.11; n = 19). Tip of genital filament in form of deep spoon, with angle of concavity highly accentuated (Fig. 1). N. neivai Holotype. Head (Table I): ratio of lengths clypeus/head 0.37 (0.41 ± 0.02; n = 9) and eyes/head 0.86 (0.89 ± 0.03; n = 9). Labrum-epipharynx (LE) long, ratio of LE/head 0.90 (0.98 ± 0.06; n = 9). Ratio between AIII/LE 1.09 (1.01 ± 0.06; n = 9). Head of holotype lying in its side, it not being possible to measure the interocular distance. Ratio clypeus/interocular distance in remaining insects 1.00 ± 0.06 (n = 9). Antennal formula AIII-AXIII 2; AXIV-AXVI 0. Ascoids long, their apex being close to the end of the flagellomere. Internal and external ascoids implanted at the same level. Pre-apical papilla present on AIII, AIV AXIV, AXV and AXVI. Simple bristles absent from AIII to AXIII. Two final antennal segments of variable size, AXV being equal to AXVI (n = 1) or shorter (n = 4). Newstead's spines of 3rd palpal segment grouped on median third in area wider than rest of segment. Palpal formula 1.4.2.5.3, that in the remaining specimens being 1.4.2.3.5 (n = 9). Cervix: ventro-cervical sensilla absent. Thorax (Table II): notum and paratergite dark chestnut, pleurae pale. Proepimeral and superior anepisternal bristles present. Anterior region of katepisternum without bristles. Wings measuring 600 (524 ± 18; n = 9) long, R5 1348 (1133 ± 27; n = 9) wide. Principal wing indices: a (alpha) 544 (476 ±26; n = 9); b (beta) 283 (247 ± 18; n = 9); g (gamma) 238 (188 ± 18; n = 9); d (delta) 295 (240 ± 27; n = 9). Tarsomeres III and IV with thin spines, implanted at two levels, one apical and the other median. Anterior leg lost in the holotype, anterior, median and posterior femur in the other examples measuring 643 ± 22 (n = 8), 680 (623 ± 24; n = 7), and 725 (693 + 26; n = 7) respectively. Tibia of anterior, median and posterior leg measuring 762 + 21 (n = 8), 1042 (934 ± 24; n = 7), and 1258 (1139 ± 56; n = 6) respectively. Tarsomere I of anterior, median and posterior leg 412 ± 12 (n = 8), 589 (499 ± 26; n = 7) and 680 (574 ± 25; n = 6) respectively. Abdomen (Table III): tergal papillae absent. Gonocoxite: 289 (279 ± 13; n = 9) long by 50 (64 ± 5; n = 9) wide. Basal tuft of bristles absent, only few sparse bristles in median area. Gonostyle measuring 139 (134 ± 5; n = 9) long, presenting four strong spines, one being apical, one external superior implanted on the apical fourth, one external and one internal. Latter two are implanted at same level, on the apical half of the structure. Pre-apical bristle absent. Paramere digitiform, wide at base and with a sharp edge to ventral region, with groups of fine bristles in the dorsal margin of the apical dorsal region and ventral margin, implanted on the widest part of the paramere. Lateral lobe 269 (267 ± 12; n = 9) long, with ratio gonocoxite/lateral lobe 1.07 (1.05 ± 0.07; n = 9). Aedeagus conical. Ejaculatory pump with wide body. Genital filament short, ratio genital filament/ejaculatory pump 1.60 (1.49 ± 0.08; n = 9). Tip of genital filament distended, in form of shallow spoon (Fig. 2). Redescription of females of N. intermedia and N. neivai N. intermedia Head (Table IV): ratio between lengths of clypeus/head 0.49 (0.48 ± 0.04; n = 19), and eyes/head 0.88 (0.88 ± 0.03; n = 19). Labro-epipharynx long, ratio labro-epipharynx/head 1.36 (1.37 ± 0.07; n = 19) and AIII/labro-epipharynx 0.65 (0.64 ± 0.03; n = 19). Ascoids implanted at same level, without basal prolongation basal, distal prolongation being long, reaching apex of subsequent segment. Antennal formula AIII-AXIII 2; AXIV-AXVI 0. Simple bristles absent from AIII to AXIII. AXV shorter than AXVI (n = 11), may be equal (n = 3) or shorter (n = 2). Palpal formula highly variable, most common being 1.4.5.2.3 (n = 11), but 1.4.2.5.3 (n = 5), and 1.4.(2.5).3 (n = 4) also occurring. Newstead's spines grouped in median region of 3th palpal segment, or also implanted on apical fourth in small groups, close to each other. Lacinia of maxilla presents two longitudinal groups of 8-10 external teeth (n = 19). Cibarium with 8-10 (usually 8) horizontal teeth. Vertical teeth strong and numerous, number varying between 16 and 24. Cibarial arch complete and pigment patch well defined. Pharynx without teeth, with small rugosities in its apical region. Cervix: ventro-cervical sensilla absent. Thorax (Table V): colour and presence of bristles as in male. Wing 623 (621 ± 24; n = 19) long, R5 1394 (1350 ± 54; n = 19) long. Principal wing indices: a (alpha) 544 (557 ± 45; n = 19); b (beta) 295 (310 ± 32; n = 19); g (gamma) 192 (215 ± 21; n = 19); d (delta) 306 (298 ± 51; n = 19). Tarsome-res III and IV with thin spines, implanted at two levels, one apical and the other median. Femur of anterior, median and posterior leg measuring 702 (683 ± 26; n = 19), 714 (686 ± 26; n = 19), and 770 (750 ± 32; n =19) long respectively. Tibia of anterior, median and posterior leg measuring 816 (805 ± 40; n = 19), 1031 (1017 ± 45; n = 18), and 1280 (1252 ± 75; n = 19) long respectively. Tarsomere I of anterior, median and posterior leg measuring 442 (429 ± 24; n = 19), 555 (548 ± 31; n = 18), and 634 (624 ± 33; n = 19) long, respectively. Abdomen (Table VI): spermathecae annulated (Fig. 3), measuring 47 (56 ± 6; n = 19) long by 17 (17 ± 1; n = 19) wide (Fig. 3). Number of annulations varying between 8-15, usually 10 or 11. Sum of annulations in two spermathecae varying from 17-33, usually 22. Head of sper-matheca about 11 (11 ± 2; n = 19) long, 12 (12 ± 2; n = 19) broad at its apex and 10 (10 ± 2; n = 19) at base, simple form predominating but simple/bilobed and bilobed forms also occurring. Individual ducts striated, with some excrescences in its insertion with the spermatheca, measuring 97 (99 ± 12; n = 19) long. Common duct 47 (48 ± 10; n = 19) long (Fig. 4). Ratio individual/common ducts 2.06, varying between 1.20 and 3.05 (2.14 ± 0,52; n = 19) (Fig. 5). Bristles without special characteristics, measuring 144 (144 ± 10; n = 19) long. N. neivai Head (Table IV): ratio between lengths of clypeus and head 0.47 (0.46 ± 0.03; n = 9), eyes/head 0.91 (0.88 ± 0.02; n = 9). Labro-epipharynx long, ratio labro-epipharynx/head 1.31 (1.31 ± 0.09; n = 9) and AIII/labro-epipharynx 0.67 (0.64 ± 0.04; n = 9). Ascoids implanted at same level in all segments, without presenting basal prolongation, apical prolongation reaching to apex of next segment. Antennal formula AIII-AXIII 2; AXIV-AXVI 0. Simple bristles absent on AIII-AXIII; pre-apical papillae present on AIII, AIV, AXIV, AXV and AXVI. AXV shorter than AXVI (n = 4). Two palpal formulae encountered, i.e., 1.4.5.2.3 (n = 7) and 1.4.(2.5).3 (n = 2). Newstead's spines found dispersed over median region of 3rd palpomere, or in two or three small groups. Lacinea of maxilla presenting two longitudinal groups of 7-10 external teeth (n = 9). Cibarium with 8-10 horizontal teeth, usually 10. Vertical teeth strong, number varying between 17-24. Cibarial arch complete and pigment patch well defined. Pharynx without teeth, with small rugosities in apical region. Cervix: ventro-cervical sensilla absent. Thorax (Table V): colour and presence of bristles as in male. Wings measuring 567 (633 ± 22; n = 8) wide, R5 1178 (1321 ± 15; n = 8) long. Principal wing indices: a (alpha) 465 (579 ± 35; n = 8); b (beta) 329 (285 ± 18; n = 8); g (gamma) 204 (224 ± 21; n = 8); d (delta) 215 (313 ± 16; n= 8). Tarsomeres III and IV with thin spines, implanted at two levels, one apical and other median. Femur of anterior, median and posterior leg measuring 634 (697 ± 20; n = 6), 634 (691 ± 14; n = 5) and 748 (759 ± 19; n = 8) long respectively. Tibia of anterior, median and posterior leg measuring 668 (780 ± 15; n = 6), 884 (997 ± 37; n = 5), and 1190 (1215 ± 28; n = 8) long respectively. Tarsomere I of anterior, median and posterior leg measuring 363 (440 ± 6; n = 4), 442 (525 ± 20; n = 4), and 589 (600 ± 14; n = 7) long, respectively. Abdomen (Table VI): spermatheca formed by 7-10 (usually 8 or 9) annulations (Fig 4), measuring 39 (42 ± 3; n = 10) long by 14 (14 ± 1; n = 10) wide (Fig. 5). Sum of annulations of two spermathecae varying between 13-22, usually 17. Head of spermatheca simple in the majority of examples, measuring 8 (8 ± 1; n = 10) long and 8 wide (8 ± 1; n = 10) at its apex, and six (6 ± 1; n = 18) at its base. Individual ducts lightly striated, measuring 64 (71 ± 11; n = 6) long. Common duct 22 (22 ± 5; n = 5) long (Fig. 6). Ratio individual/common duct 2.91, varying from 2.68-4.41, (3.42 ± 0.73; n = 5) (Fig. 6). Cercae without special characteristics, measuring 152 (148 ± 11; n = 10) long. MATERIAL EXAMINED N. intermedia - Material collected at Além Paraíba, Minas Gerais by JD Andrade Filho, with Falcão light trap, on 27/08/2002, deposited in the collection of the Centro de Pesquisas René Rachou-Fiocruz, total number still not determined, with entry number 3143/02. N. neivai - Holotype male, collected at Instituto Butantan, São Paulo, by R Fischer, on 02/07/1926, deposited in collection of Instituto Oswaldo Cruz-Fiocruz, with accession number 2267. The other examples used were laboratory-reared examples of this species (F2), collected in the municipality of Conchal, São Paulo, by C Casanova. DISCUSSION Based on morphological characteristics such as absence of ventro-cervical sensilla, and pilosity in the anterior region of the catepisternum, only one apical papilla present on AIII, 4th + 5th palpomeres longer than the 3rd and Newstead's spines absent on the 2nd palpomere, the two species can be placed with confidence within the genus Nyssomyia. The males of the two species can be separated from the other species of the genus by the combination of the following characters: absence of simple bristles on the basal segments of the antennae and short genital filaments, the genital filament/ejaculatory pump ratio being less than 1.60 with the body of the pump very much wider than in the other species. Females may be separated from the other species by absence of simple bristles on the basal segments of the antennae, by number of cibarial teeth, by the shape and number of annulations of the spermathecae and by the length of the ducts. The coloration of the thorax also facilitates separation of N. intermedia and N. neivai from some species. Without doubt N. intermedia and N. neivai are valid species that can be separated from each other. Although Phlebotomus mazzai Paterson, 1926 was described from material from Argentina (Paterson 1926) in the same year as N. neivai was recognized, there is no record of the date and month the description was made. According to the International Code for Zoological Nomenclature, chapter 5, article 21, when this occurs the last day of the year of publication should be taken as the description date (ICZN 1999), i.e., 31 December 1926 in this case. We thus conclude that N. neivai, whose formal description was published on 30 June 1926, is the senior synonym of P. mazzai. This synonymity also was verified by Marcondes and Lozovei (1999). No morphological features of the head or thorax were found that permitted separation of the two species. The genital filament tips were distinct, however. In N. intermedia the tip has the form of a deep spoon, while in N. neivai it is markedly shallower. The great variation found in the number of horizontal cibarial teeth do not permit separation of the females (Andrade Filho 2003). The number of spermathecal annulations may aid in the correct identification, but this feature is inadequate in the majority of cases, given that values for the two species overlap. Other aspects of this structure may be useful, however. The spermatheca of N. intermedia frequently present their maximum width in the apical annulations, diminishing gradually to the basal annulations, while in N. neivai the central annulations are normally broadest. Another important feature of the spermathecae of N. intermedia is that they may present some irregular, asymmetric annulations, which rarely occurs in N. neivai. The sum of the annulations of the spermathecae may also be useful, almost half of the examples studied presenting less than 16 annulations in N. neivai and more than 23 in N. intermedia. The shape of the head of the spermatheca may assist in the correct separation of the species, given that it is rarely bilobed in N. neivai. In addition, the spermathecal head of N. intermedia is always wider both at the apex and the base, giving it a more robust aspect than that of N. neivai. The individual and common ducts of the spermatheca may also be useful. The common duct is always longer in N. intermedia while the individual ducts are more striated than those of N. neivai and show excrescences where they join with the spermathecae. The group of characters described above or the presence of at least some of them permit females of N. intermedia and N. neivai to be separated with complete confidence. Based solely on morphology it was impossible to determine whether a third species was also present. Although unlikely to be the case, this question could be resolved using molecular techniques. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc03207t2.jpg] [oc03207f6.jpg] [oc03207f2.jpg] [oc03207t1.jpg] [oc03207f1.jpg] [oc03207f5.jpg] [oc03207t6.jpg] [oc03207t3.jpg] [oc03207f4.jpg] [oc03207f3.jpg] [oc03207t4.jpg] [oc03207t5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}