 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 98, No. 8, Nov, 2003, pp. 1083-1087 Down-modulation of Lymphoproliferation and Interferon-γ Production by β-glucan Derived from Saccharomyces cerevisiae AC Pelizon/+, R Kaneno, AMVC Soares, DA Meira*, A Sartori Departamento de
Microbiologia e Imunologia, Instituto de Biociências da Unesp *Departamento
de Doenças Tropicais e Diagnóstico por Imagem, Faculdade de
Medicina, Campus de Botucatu, 18618-000 Botucatu, SP, Brasil Financial support: Fundação de Amparo à Pesquisa do Estado de São Paulo Received 15 May
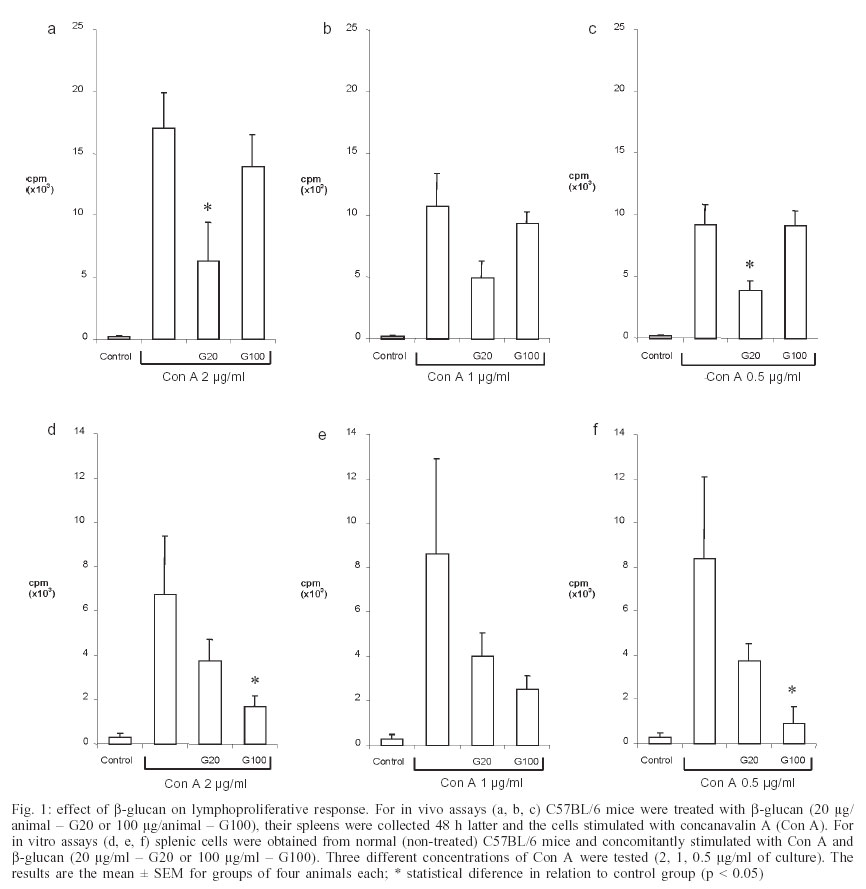
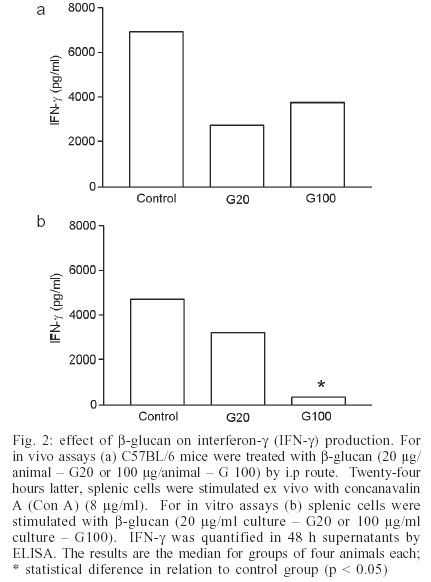
2003 Code Number: oc03211 β-glucan, one of the major cell wall components of Saccharomyces cerevisiae, has been found to enhance immu-ne functions. This study investigated in vivo and in vitro effects of β-glucan on lymphoproliferation and interferon-γ (IFN-γ) production by splenic cells from C57BL/6 female mice. All experiments were performed with particulate β-glucan derived from S. cerevisiae. Data demonstrated that both, i.p administration of particulate β-glucan (20 or 100 µg/animal) and in vitro stimulation of splenic cells (20 or 100 µg/ml of culture) decreased lymphoproliferation and IFN-γ production induced by concanavalin A. These results suggest that β-glucan can trigger a down-modulatory effect regulating a deleterious immune system hyperactivity in the presence of a strong stimulus. Key words: Saccharomyces cerevisiae - β-glucan - lymphoproliferation - interferon γ Many polysaccharides activate cellular components involved in host defense mechanisms (Tzianabos 2000). One of the most studied, β-glucan, is derived from the yeast cell walls of Saccharomyces cerevisiae and marketed as Betafectin (Masihi 2000). It affects many in vivo and in vitro activities (Estrada et al. 1997). Its main immunopharmacological activities are associated with increased host resistance to viral, bacterial, fungal, and parasitic infections (Meira et al. 1996, Sandula et al. 1999); anti-tumor activity (Di Luzio et al. 1979); radioprotectivity (Hofer & Popsil 1997); and adjuvant effect (Chirigos et al. 1978). Some animal and human investigations have failed to define the exact β-glucan action mechanism (Vetvicka et al. 1996). Some experiments have shown that it does not directly activate leukocyte functions, such as oxidative burst (Mackin et al. 1994) or cytokine secretion (Poutsiaka et al. 1993). Cytokine induction by direct interaction of glucan or zymosan with receptors on the cell surface has also been described. Glucan particles induced human monocytes to produce tumor necrosis factor-a (TNF-α) and interleukin-1b (IL-1β) by a mechanism dependent on trypsinsensitive β-glucan receptors (Abel & Czop 1992). Zymosan also induced IL-8 generation by interaction with a CD11b/CD18 receptor on human neutrophils (Au et al. 1994). More recently, vitronectin and fibronectin have been used as glucan binding proteins increasing macrophage cytokine release (Olson et al. 1996, Vassallo et al. 2001). A different view of glucan biological properties has emerged from some publications. The central idea of these reports is based on priming or sinergistic effects associated with β-glucan. Vetvicka et al. (1996) showed that soluble CR3-specific polysaccharides such as β-glucan induced a primed state of CR3 that can trigger the killing of iC3b-target cells that were otherwise resistant to cytotoxicity. Also, soluble β-1,3-glucan triggered a strong synergistic effect on IL-8, IL-10 lipopolysaccharide (LPS)-induced secretion and monocyte tissue factor activity. This glucan preparation also strongly primed LPS induced TNF-α and IL-6 (Engstad et al. 2002). A few investigations have reported down-modulatory activities mediated by β-glucan (Masihi et al. 1997, Soltys & Quinn 1999). The purpose of this study was to evaluate the effect of a non-soluble β-glucan, obtained from S. cerevisiae, on lymphoproliferation and IFN-γ production induced by conconavalin (Con A) on splenic cells from C57BL/6 female mice. MATERIALS AND METHODS Glucan - Particulate β-glucan was prepared from S. cerevisiae at the Hemocenter of Botucatu School of Medicine (São Paulo State University) using the Hassid et al. (1941) method modified by Di Luzio et al. (1979), which is briefly described here. Using a 4 l flask, 1.5 kg of dry yeast was suspended in 2 l of 3% aqueous sodium hydroxide solution. The suspension was placed in a boiling water bath for 4 h, cooled overnight, and supernatant decanted. This was repeated three times. The residue was then acidified with 800 ml concentrated hydrochloric acid (HCl) plus 2 l 3% HCl and placed in a boiling water bath for 4 h. The suspension was allowed to stand overnight, and supernatant decanted. The residue was further digested with 3 l 3% HCl at 100ºC for 4 h, cooled overnight, and decanted. The 3% HCl digestion was repeated twice. The residue was then washed with distilled water three times at 20ºC, and twice at 100ºC. Then 1 l ethyl alcohol was added to the residue, mixed thoroughly, and allowed to stand at least 24 h for maximum extraction. The particulate β-glucan preparation was then collected by filtration and dispersed with a graal homogenizer. Flasks containing 50 mg glucan were sterilized by autoclaving and a suspension was prepared for use by adding 5 ml sterile endotoxin-free saline. Animals - Female four to six-week-old C57BL/6 mice were used. They were maintained in the animal facility of the Department of Microbiology and Immunology at the Institute of Biosciences, São Paulo State University. Drug treatment - For in vivo assays the animals were treated with β-glucan (20 or 100 µg/mice) by i.p route. Spleens were aseptically obtained 48 h later and splenic cells stimulated with Con A (Sigma Chemical Co, St. Louis, MO, US) to evaluate lymphoproliferation and IFN-γ production. Animals injected with endotoxin-free saline were used as controls. For in vitro assays, splenic cells from normal mice were simultaneously stimulated with glucan and Con A. Lymphoproliferative response assay - Splenic cells were washed and adjusted to 4x106 cells/ml in complete medium (RPMI 1640 medium, Sigma) supplemented with 5% fetal calf serum, 20 mM glutamine, and 40 IU/l of gentamicin. The cells were cultured in 96-well flat-bottomed culture plates (Nunc, Life Tech. Inc., Maryland, MA, US) at 0.4x106 cells/well with or without Con A (2 and 0.5 µg/ml) for 72 h at 37ºC in a humidified 5% CO2 incubator. In the in vitro assays, splenic cultures were simultaneously stimulated with glucan (20 and 100 µg/ml) and Con A (2 and 0.5 µg/ml). To assess proliferation, 0.5 µCi/well of [3 H] methyl-thymidine was added during the last 18 h of culture, cells were harvested and radio-activity was measured in a scintillation counter. IFN-γ concentration measurement - Spleen cells (4x106 cells/ml) from normal or glucan-treated mice were cultured in 48-well tissue culture plates (Costar, Cambridge, MA, US) and stimulated with Con A (8 µg/ml) or medium alone in a humidified 5% CO2 incubator at 37ºC. For the in vitro assays, splenic cultures from normal animals were stimulated with β-glucan (20 or 100 µg/ml culture). After 48 h incubation, supernatants were collected and stored at 20ºC. IFN-γ concentration was later determined by a sandwich enzyme-linked immunosorbent assay (ELISA) as briefly described below. Maxisorp (Nunc, Nalge Nunc International, Denmark) was coated with capture antibody (AN18; 0.5 µg/ml, 100 µl/well) in NaHCO3 buffer (pH 9.6) by overnight incubation at 4ºC. The plate was washed with PBS containing 0.5% Tween 20 (PBST) and blocked with 5% non-fat milk at 37ºC for 1h. After washing, the plate was incubated with rIFN-γ or 50 µl of test samples at 37ºC for 1 h. The plate was washed with PBST and then treated with a biotinilated antibody (XMG-6; 0.075 µg/ml, 100 µl/well) diluted in PBST with 1% bovine serum albumin (Sigma). Subsequently, the plate was treated with avidin/peroxidase conjugate and reactions were revealed by adding OPD plus H2O2 in citrate-phosphate buffer. Color development was stopped with H2SO4 and optical density was measured at 492 nm. The monoclonal antibodies were gifts from Dr Giorgio Trinchieri (Laboratory for Immunological Research, Schering-Plough Research Institute, France). RESULTS AND DISCUSSION Fig. 1 shows the effect of β-glucan on proliferative response of splenic cells to Con A stimulation. Figs 1a, b, and c show that in vivo pretreatment with glucan inhibited proliferation. Unexpectedly, the lower dose (20 µg) triggered inhibition, whereas the higher dose (100 µg) did not interfere with proliferation. As expected, lower Con A doses (1 and 0.5 µg/ml) triggered lower proliferation rates in comparison with the higher dose (2 µg/ml). However, the same glucan effect was observed, independently of Con A concentration. Addition of β-glucan to in vitro cultures of splenic cells from non-treated animals, also inhibited Con A induced proliferative response (Fig. 1d, e, f). Even though the 20 µg dose caused a decrease in proliferation, only the 100 µg dose caused a significant inhibition of proliferative response. Splenic cells did not show any measurable proliferation when stimulated with 20 or 100 µg of β-glucan in the absence of Con A (results not shown). The production of IFN-γ induced by Con A followed a very similar pattern. Once again an apparent discrepancy was observed in relation to the dose effect. In Fig. 2a in vivo pretreatment with 20 or 100 µg of glucan both equally decreased IFN-g production; however this was not statistically significant. The in vitro addition of glucan also resulted in an inhibition of IFN-γ production. In this case inhibition was significant with the higher glucan concentration. No IFN-g was detected in cultures not stimulated with Con A. Again, β-glucan assayed in vitro in various concentrations (5, 10, 20, 50, 100 µg) did not induce IFN-γ production (results not shown). The distinctly different in vivo and in vitro inhibition profiles are hard to reconcile. It is tempting to imagine that two distinct mechanisms are involved. Maybe the in vitro phenomenon is being triggered by a glucan concentration dependent physical blockage. The in vivo results suggest that 20 and 100 µg β-glucan could have different biological consequences. A direct toxic effect on non-adherent cells by glucan or glucan associated with Con A was discarded by testing cell viability with the exclusion tripan blue test. Comparing these results with other assay reports of β-glucan effects is hampered by the diversity of protocols employed. Also most of these publications reinforce the immunoenhancing ability of β-glucan preparations (Tzianabos 2000). Variability in glucan immunomodulatory effects has also been attributed to the physical properties of the glucan preparations (Ishibashi et al. 2002). A few reports have investigated glucan effect on the proliferative response of splenic cells to polyclonal activators. Differently from our results, they have also described immuno-enhancing effects. Rios-Hernandez et al. (1994), using a non-soluble glucan preparation similar to the one in this study, found an increase in the spleen cell proliferative response to phytohemaglutinin A (PHA). Also Borosková et al. (1998), demonstrated that β-glucan derived from S. cerevisiae, administered in vivo to mice infected with Toxocara larvae, caused a marked stimulative and restorative effect on the parasite-suppressed lymphoproliferative response. Some water derivatives from particulate β-glucan were also able to increase mitogenic and comitogenic activities in vitro with PHA (Sandula et al. 1995). It is important to note that these investigations used PHA or specific antigens and not Con A as in this study. The most obvious way to explain this inhibition is a physical interaction between glucan particles and Con A, blocking mitogen binding to the T cell surface. Con A binding to glucan could occur if this preparation was contaminated with mannoproteins as described for a mannoprotein-β-glucan complex released from Candida albicans (Kurihara et al. 2003). To test this possibility, β-glucan preparation was mixed with Con A conjugated with FITC (concanavalin A FITC labelled, Sigma). This reagent gave positive results with both Candida sp. and S. cerevisiae whole yeast cells, but no fluorescence was observed when β-glucan particles were assayed by the same protocol (not shown), suggesting that our β-glucan preparation was not contaminated with mannoproteins. Another interesting possibility is suggested by Duan et al. (1994). These authors demonstrated that both zymosan and particulate β-glucan inhibited NK-cell-mediated killing of various target-cells. Zymosan also inhibited IL-2-dependent proliferation of NK cells, suggesting that some components from the yeast cell wall deliver a down-modulatory signal affecting multiple NK functions. They suggest that, by acting as a weak agonist, zymozan somehow interferes with the stronger mitogenic signal triggered by IL-2. In this context it is tempting to hypothesize that our IL-2 induced by Con A stimulated lymphocytes is, at least partially, responsible for the proliferation. Similarly to Duan et al. (1994), β-glucan could be interfering with T cell proliferation in response to IL-2. In a previous set of experiments we tested β-glucan effect on NK activity. Interestingly, β-glucan administration resulted in decreased splenic NK activity (not shown). This could be explained by NK cell migration to the β-glucan inoculation site. Maybe this in vivo β-glucan treatment is able to induce a selective migration of immunocompetent cells from the spleen to the peritoneal cavity. This could result in a lower number of Con A responsive lymphocytes in the spleen and a consequent local decrease in both IFN-g production and lym-phoproliferation. It has been demonstrated recently that β-glucan derived from C. albicans suppresses IFN-γ production by T cells in a monocyte dependent mechanism. The authors suggested induction of IL-10 as a possible inhibitory mechanism (Nakagawa et al. 2002). A similar IL-10 induction could be associated with IFN-g inhibition in our experiments. Another explanation could be that the final β-glucan effect depends on the intensity and type of other stimulus involved. In our experiment, the use of Con A as a strong stimulus associated with β-glucan, could trigger a down-modulatory activity, preventing the deleterious effect inherent to immune system over-stimulation. In our view, it is of major importance to decipher these apparently antagonic activities (stimulation x inhibition) so that better clinical use can be made of this compound. It is also important to investigate the influences of doses, extraction methodology, and sources of this immunomodulator. ACKNOWLEDGEMENTS To Izolete Thomazini for β-glucan extraction. REFERENCES
Copyright 2003 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc03211f1.jpg] [oc03211f2.jpg] |
| |||||||||

{kind=link}
{kind=link}