 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
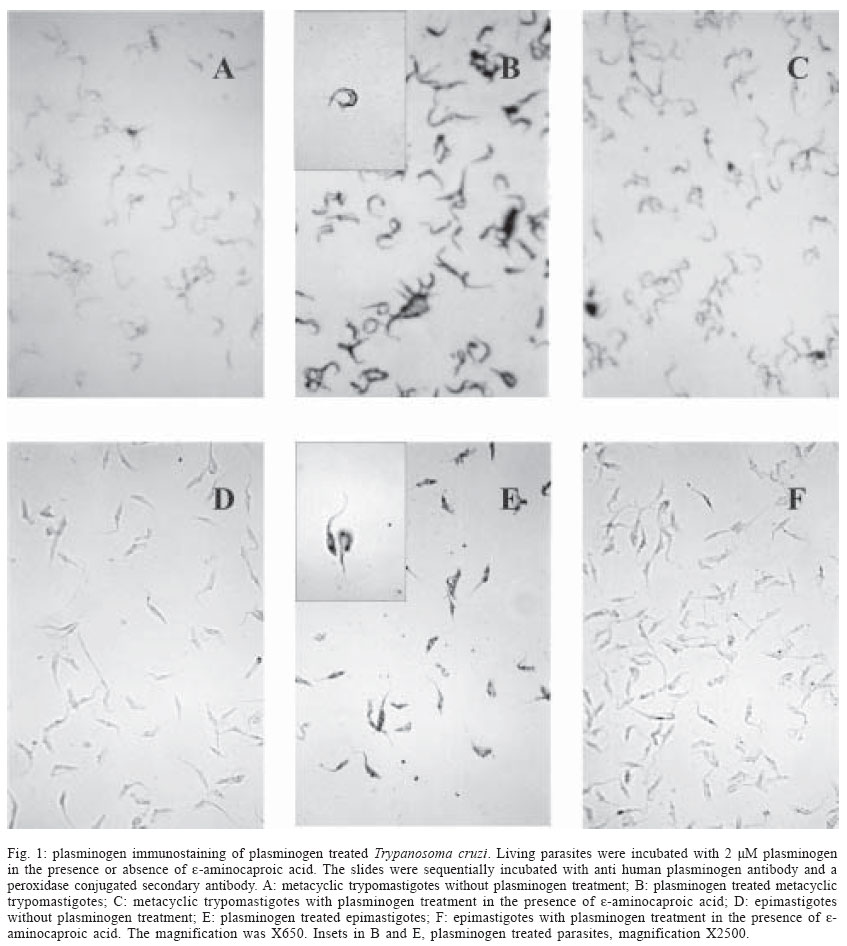
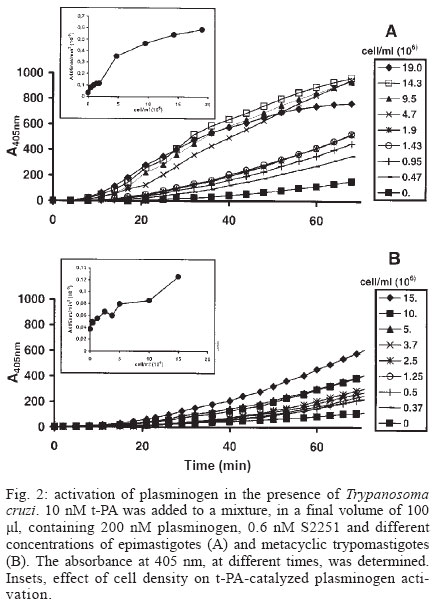
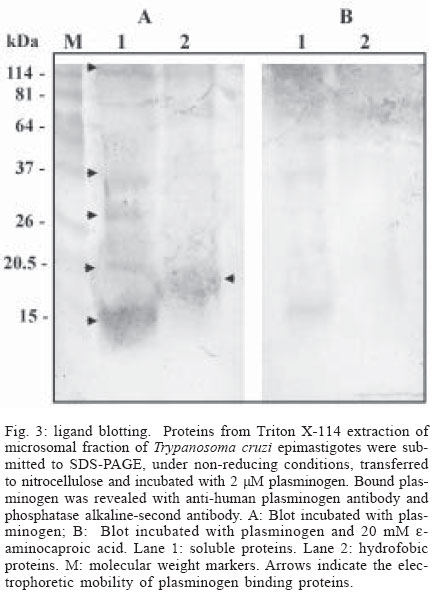
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 1, Feb, 2004, pp. 63-67 Plasminogen Interaction with Trypanosoma cruzi Laura Almeida , Gilmer Vanegas, Marina Calcagno, Juan Luis Concepción, Luisana Avilan+ Departamento de Biología, Facultad de Ciencias, Universidad de Los Andes, Mérida 5101 , Venezuela Financial support: CDCHT-Universidad de los Andes, grant C10370003B and FONACIT, grant S12001000360 Received 14 July 2003 Code Number: oc04011 ASBTRACT The ability of Trypanosoma cruzi to interact with plasminogen, the zimogenic form of the blood serin protease plasmin, was examined. Immunohistochemistry studies revealed that both forms, epimastigotes and metacyclic trypomastigotes, were able to fix plasminogen in a lysine dependant manner. This interaction was corroborated by plasminogen activation studies. Both forms of the parasite enhanced the plasminogen activation by tissue-type plasminogen activator. The maximal enhancements obtained were 15-fold and 3.4-fold with epimastigotes and metacyclic trypomastigotes, respectively, as compared to plasminogen activation in absence of cells. Ligand-blotting analysis of proteins extracted with Triton X-114 from a microsomal fraction of epimastigotes revealed at least five soluble proteins and one hydrophobic protein able to bind plasminogen. Key words: Trypanosoma cruzi - plasminogen - plasminogen activation Trypanosoma cruzi is the causative agent of Chagas disease, a public health problem in many countries of Latin America. During its life cycle, this parasite alternates between different morphological types, the epimastigote and the metacyclic trypomastigote in the insect vector and the amastigote and the bloodstream trypomastigote form in the mammal host. In the transmission from the vector to the mammal host, the parasite should transverse barriers such the extracellular matrix and reach susceptible host cells. Surface or secreted proteases have been implicated in parasite infections, among other functions, by their role in degrading proteins of the extracellular matrix (Santana et al. 1997). Additional surface protease activity for migration and spreading can be also acquired through the interaction with host proteins which do not stimulate the host's immune response. This additional activity could replace or supplement the parasite proteases. An increasing number of pathogens have the ability to bind plasminogen, a plasma and extracellular protein that is transformed, through the action of a plasminogen activator, in the broad-spectrum protease plasmin (Boyle & Lottenberg 1997, Coleman & Benach 1999). Moreover, several pathogens such as group A Streptococci, Staphylococcus aureus and Yersinia pestis, generate their own plasminogen activator (Lähteenmäki et al. 2001). Other plasminogen-binding pathogens may be capable of using host plasminogen activator (Lähteenmäki et al. 2001). In the case of Y. pestis, Borrelia burgdorferi, and group A Streptococci, the interaction with plasminogen has been demonstrated to be implicated in the invasiveness within the host (Gebbia et al. 1999, Li et al. 1999, Goguen et al. 2000). The suggested functions for plasmin in pathogenic infection include extracellular matrix protein degradation, fibrin degradation, release of peptides for nutrition, and metalloprotease activation. Since the ability to interact with plasminogen has been shown in a trypanosomatid, Leishmania mexicana (Avilan et al. 2000, Calcagno et al. 2002), we explored, in this work, the plasminogen binding ability of T. cruzi. MATERIALS AND METHODS Parasites - T. cruzi EP strain were used in this work. Epimastigote were grown in liver-infusion tryptose (LIT) medium, supplemented with 10% heat inactivated fetal bovine serum, at 28°C. Axenically derived metacyclic trypomastigotes were obtained by transferring epimastigotes in a defined medium TAU3AAG at 28°C (Contreras et al. 1994) following 3 day incubation. Metacyclic trypomastigotes were further purified using a DEAE-cellulose column as described elsewhere (De Souza 1983). Epimastigote forms used to obtain metacyclic trypomastigotes were maintained, for at least one year, by serial passage in LIT medium after being obtained from one mouse with parasitaemia. Proteins - Human plasminogen was purified to homogeneity from fresh human plasma (Deutsch & Mertz 1970). Tissue-type plasminogen activator (t-PA) was obtained from Chromogenix, Milano, Italy. The following antibodies were used for immunohistochemical studies: goat anti-human plasminogen (Calbiochem, San Diego, US) and rabbit anti-goat IgG conjugated with peroxidase (Sigma, St. Louis, US). Plasminogen immunoperoxidase staining - Living parasites (~1x107 cells) obtained under different specified conditions were incubated with 2 µM plasminogen in phosphate buffered saline (PBS), pH 7.4, containing 0.1% BSA (final volume 250 µl) for 90 min in the absence or the presence of 10 mM e-aminocaproic acid. At the end of the incubation period, the cells were pelleted, washed three times with 2 ml PBS and resuspended in the same buffer. Parasites suspension were placed onto glass slides, air dried, and fixed with acetone. Fixed parasites were sequentially incubated with normal rabbit serum, diluted 1:80, the primary goat anti-human plasminogen antibodies at 1:640 dilution, and peroxidase conjugated rabbit anti-goat IgG secondary antibodies at 1:400 dilution. A 3-amino-9-ethyl-carbazole-H2O2 substrate was used to visualize the immunoreactivity. Cells treated with normal serum and/or with the secondary antibodies alone, were used as controls. The parasite staining was evaluated by light microscopic observations. Plasminogen activation assays - Plasminogen activation rate was performed by monitoring the amidolytic activity of generated plasmin in a test volume of 100 µl. Plasminogen (200 nM) was incubated at room temperature in PBS (final volume 100 µl) containing 0.6 mM plasmin substrate S2251 (Chromogenix) in the presence or absence of parasites. Activation of plasminogen was initiated by addition of plasminogen activator (10 nM tissue-type plasminogen activator, t-PA). The hydrolysis of the chromogenic substrate was monitored continuously by measuring the absorbance at 405 nm. Initial velocities were calculated from the slope of plots A405nm versus t2 as described elsewhere (Wohl et al. 1980, Stack et al. 1991). Triton X-114 phase - Microsomal fraction were obtained by differential centrifugation from T. cruzi epimastigotes cultured in two liters of LIT medium as described previously (Caceres et al. 2003). A stock of 10% Triton X-114 was added to this microsomal fraction to a final concentration of 2% Triton X-114. Phase partition was carried out incubating the sample at -20°C overnight and then at 28°C for 10 min. Detergent (lower) and aqueous (upper) phases were recovered and the proteins precipitated with cold acetone. Ligand blotting - Microsomal proteins (soluble and hydrophobic) obtained after Triton X-114 extraction were separated by SDS-polyacrylamide electrophoresis as described by Laemmli (1970) under non-reducing condition, and parallel gels were either stained with Coomasie blue or transferred to nitrocellulose membrane as described elsewhere (Sambrook et al. 1989). For ligand-blotting, membranes were washed in PBS and blocked overnight with PBS containing 2% gelatin. The membranes were then probed with 2 µM plasminogen in PBS-gelatin for 1 h, in the presence of 2 mM phenylmethanesulphonyl fluoride (PMSF) and 2mM EDTA. In some experiments the incubation solution contained 20 mM e-aminocaproic acid. After three washings, the membrane was incubated with anti-human plasminogen antibody for an additional 1 h. Thereafter, the membrane was rinsed with PBS and incubated with the alkaline phosphatase-conjugated second antibody and revealed with 5-bromo-4-chloro-3-indolyl phosphate/nitro blue tetrazolium (BCIP/NBT) as substrate. RESULTS AND DISCUSSION A direct evidence of plasminogen binding to T. cruzi is provided by immunostaining of plasminogen-treated parasites (Fig. 1). Both metacyclics trypomastigotes and epimastigotes resulted positive to plasminogen binding (Fig.1B, 1E). Moreover, in both cases, the staining was reduced by the presence, in the incubation mixture, of e-aminocaproic acid, a lysine analogue (Fig.1C, 1F). These results suggest that the lysine binding kringle structure, homologous triple-loop structures (Ponting et al. 1992), of the plasminogen molecule might be involved in the association of plasminogen with the parasite. These lysine binding sites have been implicated in the interaction of plasminogen with fibrin and with surface proteins on a variety of cells including pathogen organisms (Angles-Cano 1994, Lähteenmäki et al. 2001). Plasminogen staining was visualized in parasites incubated in TAU3AAG medium during several days. However, the staining resulted weak with seven days of incubation (data not shown). When plasminogen interacts with molecules such as receptors or fibrin, a conformational change is produced that permit an enhancement of plasminogen activation by physiological activators (Angles-Cano 1994). To corroborate the plasminogen binding onto the T. cruzi surface, we investigated the plasminogen activation by t-PA in the presence of different cell concentrations. The experiments were performed with either epimastigotes or metacyclic trypomastigotes. As shown in Fig. 2, both types of forms were able to facilitate the plasminogen activation by t-PA in cell concentration-dependent manner, the epimastigote form being more effective in producing the plasminogen activation enhancement. The maximal enhancements obtained were 15 ± 3.2 and 3.4 ± 1 for epimastigotes and metacyclic trypomastigotes, respectively, compared to plasminogen activation in absence of cells. No plasmin activity was observed when plasminogen activator was omitted (data not shown) indicating that T. cruzi cells do not produce a plasminogen activator. The plasminogen activation enhancement by T. cruzi shows that this parasite is able to bind plasminogen on its surface and that T. cruzi can act as a regulator of this activation. The lower enhancement of plasminogen activation by metacyclic trypomastigotes compared to epimastigotes could reflect the molecular changes associated to culture conditions. Axenic metacyclic forms derived from epimastigotes maintained more than 78 days in culture are less virulent and have different protein profile than the insect-born metacyclic trypomastigotes (Contreras et al. 1998). By contrast, the protein profile of epimastigotes do not change by several passage in axenic medium (Contreras et al. 1998). Finally, an attempt to identify T. cruzi plasminogen receptor was performed using a ligand-blotting assay on proteins from a microsomal fraction extracted with Triton X-114. The treatment with this detergent allows to separate hydrophilic proteins (aqueous phase) and membrane bound proteins (detergent phase). This assay allowed the demonstration of at least five plasminogen binding proteins in the soluble fraction, of 110, 37, 32, 19, and 13 kDa of molecular masses and one plasminogen binding protein of 15 kDa of molecular mass in the detergent fraction (Fig. 3A). Control blotting in which plasminogen was omitted showed no band reactivity (data not shown). Incubation of membranes with plasminogen and e-aminocaproic acid abolished the binding to the parasite proteins, indicating specific binding through lysine binding sites in the plasminogen molecule (Fig. 3B). Several proteins have been identified as plasminogen receptors in numerous cell types such as vertebrate cells and pathogenic organisms (Plow et al. 1991, Lähteenmäki et al. 2001, Crowe et al. 2003). Moreover, in Candida albicans, at least six plasminogen binding proteins were found (Crowe et al. 2003). These receptors are characterized by a relative low affinity for plasminogen. The identification of the plasminogen binding protein in T. cruzi will allow determining whether it is structurally related to other proteins which have been identified as plasminogen receptor. The interaction with plasminogen may be important in the establishment of the infection into the vertebrate host where the plasminogen is available. Plasminogen activated on the parasite surface could contribute to cross barriers for migration. The fact that metacyclic trypomastigote is able to bind plasminogen is consistent with this hypothesis. However, in this study we found that epimastigote, a form that is exclusively found in the vector host, is also able to interact with plasminogen. Moreover, this form produced a higher enhancement of plasminogen activation than the axenic metacyclic trypomastigote. One may speculate that plasminogen-binding molecules could have additional functions in the parasites and for that the plasminogen binding property may be found in vitro in either epimastigotes or in metacyclic trypomastigote. Different functions for plasminogen receptors have been found in pathogens. In group A streptococci, the two active enzymes, glyceraldehyde 3-phosphate dehydrogenase and a-enolase, both found on cell surface, are also plasminogen binding molecules (Lähteenmäki et al. 2001). Another possible explanation for the presence of plasminogen receptor in epimastigote is that plasminogen provided by the blood meal can have some function in the vector. Such is the case of B. burdorferi, in which the plasminogen interaction is important for the establishment in the vector (Coleman et al. 1997). To know whether the plasminogen interaction with T. cruzi has a physiological function in either the vertebrate host or the insect vector, in vivo studies are required such as experimental infection in plasminogen-deficient transgenic mice and vector-feeding experiments. ACKNOWLEDGMENTS To Dr Victor Contreras for help with the metacyclic trypomastigote culture. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc04011f3.jpg] [oc04011f2.jpg] [oc04011f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}