 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
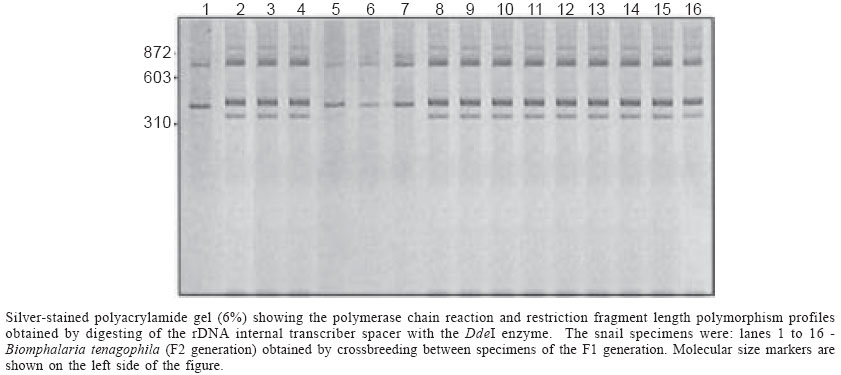
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 1, Feb, 2004, pp. 85-87 Dominant Character of the Molecular Marker of a Biomphalaria tenagophila Strain (Mollusca: Planorbidae) Resistant to Schistosoma mansoni Florence Mara Rosa, Roberta Lima Caldeira*, Omar dos Santos Carvalho*, Ana Lúcia Brunialti Godard***, Paulo Marcos Zech Coelho**/+ Departamento de Parasitologia ***Departamento de Biologia Geral, Instituto de Ciências Biológicas-UFMG, Belo Horizonte, MG, Brasil *Laboratório de Helmintoses Intestinais **Laboratório de Esquistossomose, Centro de Pesquisas René Rachou-Fiocruz, Av. Augusto de Lima 1715, 30190-002 Belo Horizonte, MG, Brasil Partial financial support: CNPq-Pronex and Fapemig, Brazil Received 18 August 2003 Code Number: oc04015 ABSTRACT Biomphalaria tenagophila population from Taim (state of Rio Grande do Sul, Brazil) is totally resistant to Schistosoma mansoni, and presents a molecular marker of 350 bp by polymerase chain reaction and restriction fragment length polymorphism of the entire rDNA internal transcriber spacer. The scope of this work was to determine the heritage pattern of this marker. A series of cross-breedings between B. tenagophila from Taim (resistant) and B. tenagophila from Joinville, state of Santa Catarina (susceptible) was carried out, and their descendants F1 and F2 were submitted to this technique. It was possible to demonstrate that the specific fragment from Taim is endowed with dominant character, since the obtained segregation was typically mendelian. Key words: Biomphalaria tenagophila - polymerase chain reaction and restriction fragment length polymorphism - Biomphalaria tenagophila is a planorbid with a wide distribution in South America (Paraense 1984), and has epidemiological importance, since this species maintains the cycle of the trematode Schistosoma mansoni in some areas in Brazil. In spite of its dominance in some areas, B. tenagophila is found in the nature with low rates of infection, however, it is responsible for the majority of the autochthonous cases of schistosomiasis in the state of São Paulo, as well as for the foci of the disease in the states of Minas Gerais and Santa Catarina (Paraense 1986). This last focus is maintained by a highly susceptible population (Joinville) of molluscs to S. mansoni. On the other hand, there is a population coming from the Ecological Reservoir from Taim (state of Rio Grande do Sul), that has systematically been found resistant to different strains of S. mansoni (Coelho 1995). This population has been maintained under laboratory conditions for more than twenty years, without any selective pressure favoring the resistance. Vidigal et al. (1998), using the polymerase chain reaction and restriction fragment length polymorphism (PCR-RFLP) of the ribosomal RNA internal transcribed spacer region (ITS) with restriction DdeI enzyme, were able to separate molecularly the molluscs genus Biomphalaria, such as B. glabrata, B. tenagophila, and B. straminea, which are intermediate hosts of S. mansoni in Brazil. Those authors obtained species-specific profiles for this three species, observing besides that various Brazilian populations of B. tenagophila showed a characteristic profile, with two fragments (800 and 470 pb). Barbosa (2001), using the same technique, observed that the Taim population, in addition to the characteristic species-specific profile of B. tenagophila, as described by Vidigal et al. (1998), has a 350 pb fragment too. In the present study, cross-breedings between molluscs from B. tenagophila (Taim) and B. tenagophila (Joinville) were carried out, aiming at verifying whether the marker of the Taim population has a dominant character. Studies in order to obtain F1 generation were undertaken using a specimen of B. tenagophila Taim and another one of albino B. tenagophila Joinville. They were maintained together in order to achieve cross-breeding (4 pairs). The albino recessive character was used, in this case, as a phenotypic marker. The molluscs with 5 mm in diameter, approximately, sexually immature, were put together into a plastic recipient with 100 ml dechlorinated water for 50 days. Afterwards, the respective pairs were separately put into glasses in order to obtain F1 descendants. In this experiment, the egg-layings from the albino snails, as well as from the pigmented ones, were collected for four weeks. The egg-layings from F1 generation were transferred to an aquarium until the molluscs could reach 5 mm in diameter. F2 generation was obtained by means of cross-breedings between F1 individuals according to the same procedures previously described. A fragment from the cephalopodal region of the molluscs was collected for DNA extraction, using the Wizard Genomic DNA Purification Kit (Promega), as modified by Vidigal et al. (2000). Ten specimens from Taim and 10 from Joinville populations, as well as 9 individuals from F1 generation and 16 from F2 generation were aleatory selected and were submitted to PCR-RFLP analysis. The entire ITS region was amplified using the primers ETTS2 (5-TAACAAGGTTTCCGTAGGTGAA-3) and ETTS1 (5-TGCTTAAGTTCAGCGGGT-3) (Kane & Rollinson 1994). PCR-RFLP conditions were the same described by Vidigal et al. (1998), with DdeI enzyme. Cross-breeding between Taim and Joinville populations was initially confirmed by means of a phenotypic marker (utilization of an albino lineage) and a genotypic one (350 pb fragment), which has been observed only in Taim population up to now. The specimens of F1 generation presented with pigmentation, whereas in F2 generation there were pigmented and albino individuals. The pioneer study on the use of albinism in molluscs was carried out by Boycott et al. (1929), dealing with the inheritance of sinistrality in Lymnaea peregra. In this case, the recessive albino character was used as phenotypic marker, which allowed to distinguish between the individuals generated from cross-breeding and those ones generated from self-fertilization. Posteriorly, Paraense (1955) used the albino lineage aiming at observing the reproductive behavior in Biomphalaria. These studies demonstrated that the molluscs are able to perform self-fertilization, but in the presence of one or more individuals they prefer cross-breeding. DNA amplification with the ETTS1 and ETTS2 primers generated a fragment of approximately 1300 bp for all specimens. The Figure shows the profile of B. tenagophila obtained after restriction of this fragment by the DdeI enzyme. The Taim population showed a profile with three fragments (800, 470, and 350 pb), the Joinville population two fragments (800 and 470 pb) and the F1 generation, obtained by cross-breeding between Taim and Joinville populations, presented with three fragments (800, 470, and 350 pb) (data not shown). F2 generation obtained by means of cross-breeding between F1 individuals showed 12 specimens with three fragments and four other ones with two fragments (Figure). The sum of sizes of the bands produced upon digestion was not equal to the size of the undigested PCR products for Taim populations. It might be due to additional bands. As there are many copies of rDNA cistrons in the genome, we believe that such variation could be due to heterogeneity, i.e. the presence of two or more sequence types of the ITS in a single organism caused by insertion/deletion event(s) occurring outside this sequenced region or there is another restriction site(s) located very close to the extremities of this PCR product. These results demonstrated that segregation of the 350 pb fragment was typically mendelian (F2 with 3:1 proportion). This band is a genetic marker of great importance, since albino populations are rarely seen in the nature, thus being difficult to study cross-breedings among populations. It is speculated that the specific marker of the Taim population will be important in monitoring a possible biological control, which consists in placing specimens from Taim together with the susceptible local population, in order to obtain a possible crossbreeding between both populations, thus fixing the resistance character of the mollusc to infection since the resistance gene has dominant character too (Rosa 2002). Briefly, even though the 350 pb fragment has no relation to resistance in these molluscs, since the region of the genoma studied is the ribosomal RNA gene intergenic spacer, that fragment will be of great value in assessing whether the genetic characters of Taim population can be inherited by future generations. ACKNOWLEDGEMENTS To Dr W Lobato Paraense for providing the albino strain of Biomphalaria tenagophila used in this work. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. Free, full-text also available from http://www.memorias.ioc.fiocruz.br The following images related to this document are available:Photo images[oc04015f1.jpg] |
| |||||||||

{kind=link}