 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 2, March, 2004, pp. 223-226 SHORT COMMUNICATION Ivermectin Resistant and Susceptible Third-stage Larvae of Haemonchus contortus: Cholinesterase and Phosphatase Activities Consuelo Giménez-Pardo/+, Maria Mercedes Martínez-Grueiro*, Alicia Gómez-Barrio*, Filomena Rodríguez-Caabeiro Departamento de
Microbiología y Parasitología, Facultad de Farmacia, Universidad
de Alcalá, Crtra. Madrid-Barcelona km 33, 28871 Alcalá de Henares,
Madrid, España *Departamento de Parasitología, Facultad de
Farmacia, Universidad Complutense, 28040 Madrid, España Received 24 June
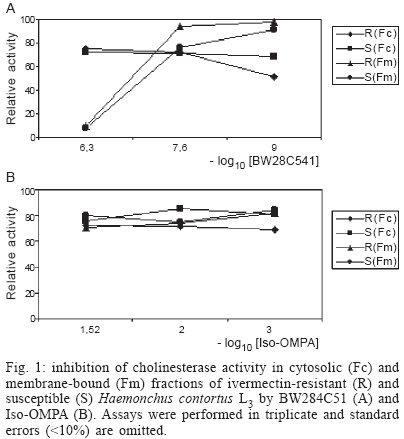
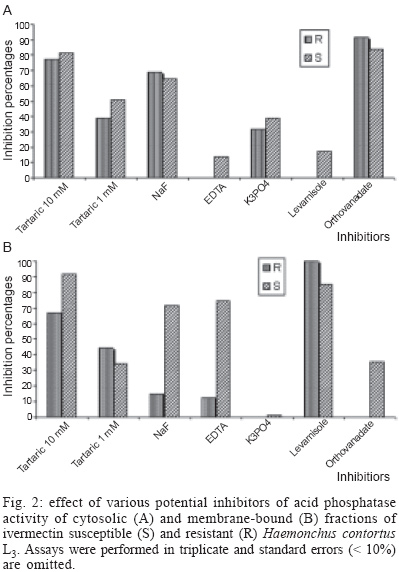
2003 Code Number: oc04040 Cholinesterase and acid phosphatase (AP), but not alkaline phosphatase activities, were detected in cytosolic and membrane-bound fractions of ivermectin resistant and susceptible Haemonchus contortus infective-stage larvae. Some differences in acetylcholinesterase activity of cytosolic fractions and in the AP activity of these fractions as well as in the response to AP inhibitors by membrane-bound fractions were detected. Data are discussed. Key words: Haemonchus contortus - cholinesterase - phosphatase - ivermectin - resistant It is known that parasite enzymatic activities are very variable among different species, sexes, and developmental stages of the biological cycle of the nematodes (Sanderson & Ogilvie 1971, Rhoads 1981, Lawrence & Pritchard 1993, Alonso-Villalobos & Martínez-Grueiro 2000, Giménez-Pardo et al. 2003). Recent studies have also revealed differences among isolates resistant and susceptible to several antihelmintics i.e. morantel tartrate (Sutherland 1987), ivermectin (Echevarría et al. 1992) benzimidazoles (Sutherland & Lee 1993) or more recently to levamisole (Giménez-Pardo et al. 2003). Some authors have suggested that after treatment with some antihelmintics, nematodes have a reduced metabolism which may have an effect on production of enzymatic activity by these nematodes (Sharpe & Lee 1981). Others have observed differences in propionyl esterases between ivermectin-resistant (IVM-R) and ivermectin-susceptible (IVM-S) strains of Haemonchus contortus (Echevarría et al. 1992). In the present paper we have studied the possibility that hydrolases, like cholinesterases and phosphatases, that play important biological roles (Massoulié et al. 1993, Selkirk et al. 2001, Martínez-Grueiro 2002) would also express differences in an IVM-R isolate and an IVM-S isolate of H. contortus L3 larvae. Both IVM-R and IVM-S isolates of H. contortus were obtained as frozen samples from the Institute for Animal Science and Health (ID-DLO), Lelystad, The Netherlands. The organization which supplied the nematode periodically checked the strains for susceptibility or resistance to ivermectin. After thawing, the helminths were resuspended in an 10 mM Tris-HCl buffer pH 7.2, with penicillin (1 mg/ml) and dehydrostreptomycin sulphate (2 mg/ml), homogenized in a glass Potter-Elvejem homogenizer and samples ultracentrifugued at 100,000 x g in a Beckman XL-100 ultracentrifugue for 30 min at 4ºC. Supernatants were centrifuged under the same conditions: this second supernatant was kept as the cytosolic fraction. To obtain the membrane-bound fraction, homogenates were prepared as mentioned above, and the pellet was suspended in a 1 ml of 10 mM sodium citrate buffer pH 6.5 containing 1 ml of 0.2 (v/v) Triton X-100 for 3 h with frequent stirring on a vortex (Heidolph, Reax Top) for 1 min every 30 min. Each Triton extract was then centrifuged as before. The supernatant was saved and the pellet was extracted again for an additional 3 h. The Triton X-100 supernatants were pooled as the membrane-bound fractions. When required, Triton X-100 was removed from the preparations by the addition of 0.3 g SM-2 Bio Beads (Bio Rad) per mililiter of the preparation and shaking gently for 2 h at 4ºC (Chang & Opperman 1991). Finally, samples were aliquoted and conserved at -80 ºC until use. Protein amounts of the cytosolic fractions was 1.47 mg/ml for the IVM-R isolate and 1.37 mg/ml for the IVM-S isolate as determined by the Bradford method (Bradford 1976). Due to the potential interaction of the Coomasie contained in Bradford reactive with the detergent (Kirazov et al. 1993), protein amounts for the membrane-bound fractions were determined by the Peterson method (Peterson 1983), and was found to be 2.28 mg/ml for the IVM-R isolate and 2.31 mg/ml for the IVM-S isolate. Cholinesterase (ChE) activity and phosphate activities were determined by a modified Ellman test (Ellman et al. 1961, Rathaur et al. 1987) and Moulay and Robert-Gero (1995) method, respectively. Quantification of the ChE activity was performed using acetylthiocholine iodide (ATCI) as substrate. The increases in OD were converted to units per litre. One unit was the equivalent to 1 nmol of substrate hydrolized per min per mg of protein and results are expressed as the mean ± standard error of three independant determinations. In the inhibition assays (20 µl-samples of cytosolic or membrane-bound fraction) plus 5 µl of each inhibitor were first incubated at 22ºC for 10 min before measuring the increase in OD. The final concentrations and inhibitors used were: 500 nM, 25 nM, and 1nM of 1,5-bis(4-allyl(dimethyammonium phenyl)pentan 3 one dibromide) (BW2845C51) and 30 mM, 10 mM and 1 mM of tetraisopropyl pyrophosphoramide (iso-OMPA). Quantification of the phosphatase activity of cytosolic and membrane-bound fractions was performed using 10 mM p-nitrophenyl phosphate (pNPP) (Sigma, St. Louis, MO, US) as substrate and the reaction product was measured at 405 nm, using an Ultrospec III Spectrophotometer (Pharmacia, LKB). One unit of specific activity was defined as 1 µmol of p-nitrophenol hydrolized per hour per mg of protein under assay conditions (Giménez-Pardo et al. 2003). Determinations were carried out with 50 mM sodium acetate buffer at several pH values (3.5-6.0), 50 mM Tris-HCl (pH 7.0-9.0) and 50 mM diethanolamine (pH 10.0-11.5). Phosphatase from different samples was also assayed in the presence of several phosphatase inhibitors to their optimum pH values. Inhibitors and their final concentrations were: 10 mM and 1 mM of L(+) tartaric acid, 1 mM NaF, 10 mM ethylenediaminetetraacetic acid (EDTA), 10 mM monobasic potassium phosphate, 10 mM levamisole, 10 µM okadaic acid and 100 µM sodium orthovanadate. Inhibitors were preincubated with the samples in the reaction buffer at 22ºC for 10 min before the addition of the substrate. Appropriate media and reagent blanks were run in parallel and test values corrected using these blanks. Cytosolic ChE activity was six times higher in the IVM-R isolate (12.3 ± 0.7 nmol min-1 mg-1) than in the IVM-S isolate (2.0 ± 0.1 nmol min-1 mg-1). Although this difference was very notable, we have previously detected in other H. contortus isolates (susceptible and resistant to levamisole) (Giménez-Pardo et al. 2003) a ChE activity similar to the detected activity in the IVM-R isolate analyzed in this study. If the difference now observed is real or not requires further studies. As in benzimidazole-resistant and susceptible isolates (Sutherland & Lee 1993), the raised levels of ChE of the resistant isolate could be related to antihelmintic resistance. Krause et al. (1998) suggested that ivermectin could act as a positive allosteric effector of the alpha7 neuronal nicotinic acetylcholine receptor. Values of 23.3 ± 4.0 nmol min-1 mg-1 for the IVM-R isolate and 20.4 ± 3.5 nmol min-1 mg-1 for the IVM-S isolate were obtained in the membrane-bound fractions after the detergent was removed (Tarrab-Hazdai et al. 1984). It is also remarkable that in the membrane-bound fractions, activities were very similar to that previously reported (Giménez-Pardo et al. 2003). ChE may also be distinguished by their affinity for, or reactivity with various selective inhibitors: BW284C51 for acetylcholinesterases and iso-OMPA for pseudocholinesterases. Results have revealed that enzymatic activity is a "true" acetylcholinesterase in both IVM-R and IVM-S isolates as both were only inhibited by the specific acetylcholinesterase inhibitor BW284C51 at the highest concentration (Fig. 1 A, B). Phosphatase activity was measured in cytosolic and membrane-bound fractions of both IVM-R and IVM-S isolates by hydrolisis of p-nitrophenylphosphate (pNPP) at several pH values (3.5-12.0). Acid phosphatase (AP) activity was detected in both isolates in the cytosolic and the membrane-bound fractions, but there was no evidence of any significative alkaline phosphatase activity in any sample. Added magnesium ions (1 mM) had no effect on the phosphatase activity at any pH value tested. Optimum pH of 5.0 for cytosolic and 4.5 for membrane-bound fractions both from IVM-R and IVM-S isolates were obtained, after Triton X-100 treatment and Bio Beads extraction. The optimum pH for each fraction falls inside the usual pH range of AP of parasitic helminths 3.0-6.0 (Maki & Yanigasawa 1980). AP activity in cytosolic fractions were 3.5 times higher in the IVM-S isolate (50.5 U) as compared to IVM-R isolate (14.9 U). By contrast, specific activity of the membrane-bound fractions was similar in both isolates (30.4 U for the IVM-R isolate and 34.5 U for the IVM-S isolate). Differences observed between cytosolic fractions are remarkable. If we compare this data with those obtained for L3 susceptible and resistant to levamisole (Giménez-Pardo et al. 2003), we observe that AP activity in IVM-R isolate is the lowest. As can be seen in Fig. 2 A, B, AP activity was also characterized with several potential inhibitors. It is well known that AP are inhibited by tartaric acid, NaF or vanadium (Beh et al. 1991). In cytosolic fractions there was no differences between the IVM-R and the IVM-S isolates and the order of potency was vanadium > tartaric acid > NaF. Little if any effect was observed with EDTA or levamisole. These observations indicate that the phosphatase activity detected corresponds to an AP. However, in the membrane-bound fractions there was differences between both isolates. In fact, the IVM-S isolate was more sensitive than the IVM-R to tartaric acid (91.9% vs 66.9%), NaF (71.5% vs 14.8%), and sodium orthovanadate (35.7% vs 0%), a specific tyrosine phosphatase inhibitor (Fischer et al. 1991). None of the fractions analyzed (cytosolic or membrane-bound) were affected by 10 µM okadaic acid, an inhibitor of serin/threonin phosphatases (data not shown). In conclusion, enzymatic activity is altered from some way in IVM-R or IVM-S isolates. However this study has been performed only on two strains, an isolate of both resistant and susceptible to ivermectin, that corresponds to an isolated geographical region. We need obviously to examinate further isolates from many different geographical regions to provide a better understanding of this fact. Experiments are now in progress. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc04040f1.jpg] [oc04040f2.jpg] |
| |||||||||

{kind=link}
{kind=link}