 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 3, May, 2004, pp.289-294 Leishmania braziliensis: Partial Control of Experimental Infection by Interleukin-12 p40 Deficient Mice Sebastião Martins de Souza-Neto, Cláudia Martins Carneiro*, Leda Quercia Vieira**/++, Luís Carlos Crocco Afonso/+ Departamento de
Ciências Biológicas, Instituto de Ciências Exatas e Biológicas,
Nupeb *Departamento de Análises Clínicas, Escola de Farmácia,
Universidade Federal de Ouro Preto, 35400-000 Ouro Preto, MG, Brasil **Departamento
de Bioquímica e Imunologia, Universidade Federal de Minas Gerais,
Belo Horizonte, MG, Brasil Financial support: Capes Received 10 November
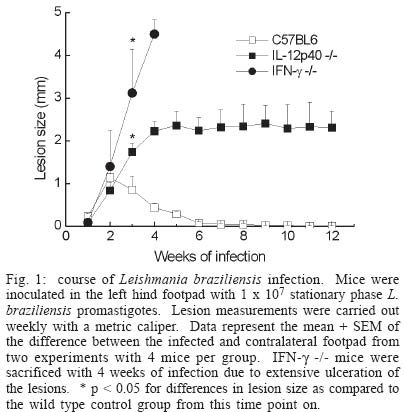
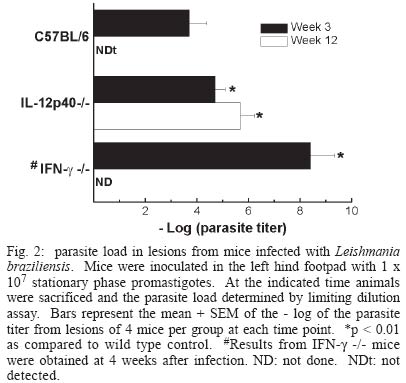
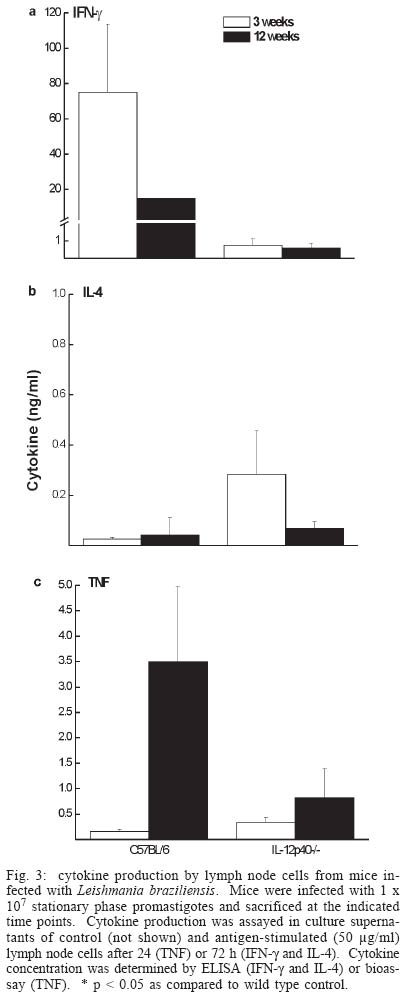
2003 Code Number: oc04052 Resistance to infection by Leishmania major has been associated with the development of a Th1 type response that is dependent on the presence of interleukin 12 (IL-12). In this work the involvement of this cytokine in the response to infection by L. braziliensis, a less virulent species in the mouse model, was evaluated. Our results show that while interferon (IFN-g) deficient (-/-) mice inoculated L. braziliensis develop severe uncontrolled lesions, chronic lesions that remained under control up to 12 weeks of infection were observed in IL-12p40 -/- mice. IL 12p40 -/- mice had fewer parasites in their lesions than IFN-g-/- mice. Lymph node cells from IL-12p40 -/- were capable of producing low but consistent levels of IFN-g suggestive of its involvement in parasite control. Furthermore, as opposed to previous reports on L. major-infected animals, no switch to a Th2 response was observed in IL-12p40 -/- infected with L. braziliensis. Key words: cutaneous leishmaniasis - Leishmania braziliensis - interleukin 12 The great majority of studies related to the immune response in cutaneous leishmaniasis using the murine model have been carried out using Leishmania major as the prototype species. These studies have established that resistance to infection by this parasite is associated with the development of Leishmania-specific Th1 cells while susceptibility and disease progression is related to a Th2 type response (Heinzel et al. 1989, 1991, Liew 1989, Scott 1989, 1991, Scott et al. 1989). While this paradigm seems to apply to most circumstances in the case of L. major, several studies have demonstrated that, especially in mice infected by other Leishmania species that also cause cutaneous leishmaniasis such as L. amazonensis, the association of susceptibility to infection and the presence of a Th2 response is not always observed (Afonso & Scott 1993, Soong et al. 1997, Jones et al. 2002). With regard to the role of cytokines in the development of the resistant phenotype, it has been demonstrated that interferon-g (IFN-g) and interleukin (IL)-12 are necessary not only in the initial steps of the infection, but also to maintain an established response against this parasite (Belosevic et al. 1989, Wang et al. 1994, Mattner et al. 1996, 1997b). The importance of IL-12 in the response to Leishmania is attributed to its involvement in the development of Th1 cells (Hsieh et al. 1993, Trinchieri 1993, Magram et al. 1996), however more recent studies have demonstrated that under certain circumstances Th1 development can be achieved in the absence of IL-12 (Scharton-Kersten et al. 1996, Brombacher et al. 1999). Several studies with murine models of L. braziliensis have demonstrated that all mouse strains tested are resistant to infection by this parasite (Childs et al. 1984, Samuelson et al. 1991, Barral-Netto et al. 1992, 1993, Lima et al. 1999) and that this resistance is dependent on IFN-g activity (DeKrey et al. 1998). Since L. braziliensis infection does not induce significant levels of IL-4, we decided to use this parasite species to further clarify the role of IL-12 in the control of infection by intracellular protozoan parasites. Our results show that, contrary to what has been observed with L. major, L. braziliensis-infected IL-12p40 -/- mice partially control parasite growth. This observation was associated with the production of low levels of IFN-g and TNF but also of IL-4, suggesting that, when infected with L. braziliensis, IL-12p40-/- mice do not default to a Th2 type response. MATERIALS AND METHODS Animals - C57BL/6 mice were obtained from Cebio (Instituto de Ciências Biológicas, UFMG, Belo Horizonte, MG, Brazil). IFN-g -/- mice were obtained from Jackson Farms (Glensville, NJ, US). IL-12p40-/- were kindly provided by Dr Luiz Vicente Rizzo from the Departamento de Imunologia, Universidade de São Paulo. Both genetically deficient strains were bred in micro isolators, given sterile food and water ad libidum. Experimental animals were males and females, 4-8 week old and were kept in a conventional animal facility. The animals were reared and used according to the Guide for the Care and Use of Laboratory Animals of the National Research Council (1996). Parasites and infection protocol - L. (Viannia) brazi-liensis promastigotes, strain M2903 (MHOM/BR/75/M2903 a gift from Dr Maria Norma Melo, Universidade Federal de Minas Gerais, were cultured in Grace's insect medium (Gibco, Grand Island, NY, US) supplemented with 20% heat-inactivated FCS (Nutricell, Campinas, SP, BR), 2 mM l-glutamine (Gibco) and 10 µg/ml gentamycin sulfate (Schering Plough, Rio de Janeiro, BR), at 26ºC. Pro-mastigotes were collected at the stationary phase (5th day of culture), centrifuged at 1540 x g at 4ºC for 20 min and washed in phosphate buffered saline (PBS). Mice were inoculated with 1 x 107 or 1 x 105 stationary phase promastigotes (5th day of culture) in the left hind footpad. Lesion development was measured during the course of infection with a dial micrometer model 1015MA (LS Starret Co., Itu, SP, Brazil) and expressed as the difference in size between the infected footpad and the contralateral uninfected footpad. Antigen preparation - Leishmania antigen was obtained from logarithmic phase cultures of L. braziliensis promastigotes. Promastigotes were washed twice in PBS and pellets were stored frozen at 20ºC until enough material was accumulated for preparation of antigen. Pellets were submitted to seven cycles of freezing in liquid nitrogen followed by thawing at 37ºC. The preparations were visually inspected for the presence of intact parasites. Protein content of preparations was assayed by the Lowry method (Lowry et al. 1951) and adjusted to 1 mg protein/ml. Antigen preparation was aliquoted and stored frozen at -70ºC. Antigens were thawed immediately before use in cell cultures. Estimation of parasite load - The number of parasites in the footpad was estimated by limiting dilution assay. Mice were sacrificed and the whole lesion was removed and weighed. A fragment of known weight was obtained and ground in cold PBS containing 10 µg/ml gentamycin (Schering Plough), in glass tissue grinders. Tissue debris were removed by centrifugation at 50 x g for 1 min, the supernatants were transferred to another tube and centrifuged at 1540 x g for 15 min. The pellet was resuspended in 0.5 ml of Grace's medium containing 20% FCS, 2 mM l-glutamine and 10 µg/ml gentamycin. The parasite suspension was then serially diluted in 10-fold dilutions in duplicates in 200 µl final volume in 96-well plates. Pipet tips were replaced for each dilution. Plates were incubated for 10 days at 26ºC and examined under an inverted microscope. Results are expressed as the negative logarithm of the last dilution in which parasites were detected, adjusted for the weight of the tissue sample. Analysis of cytokine production - Single-cell suspensions were prepared from the popliteal lymph nodes of the infected footpad harvested at 3 or 12 weeks of infection. Cells were adjusted to a concentration of 5 x 106/ml in Dulbecco's minimal essential medium (Gibco) containing 10% fetal calf serum (Nutricell), 2 mM l-glutamine and 10 µg/ml gentamycin (Schering Plough) 25 mM N-2-hydroxiethylpiperazine-N'-2-ethanosulfonic acid (HEPES) and 50 µM 2-mercaptoethanol, and plated at 0.5 ml per well in 48-well tissue culture plates. Parasite antigen was added at the start of the culture (50 µg/ml). Supernatants were harvested after 24 h for tumor necrosis factor (TNF) measurement or 72 h for IFN-g and IL 4 determination. IFN-g was assayed in a two-site ELISA using a rat anti-IFN-g monoclonal antibody R46A2 (ATCC) and a polyclonal rabbit serum specific for the cytokine, kind gift from Dr Jay Farrell, University of Pennsylvania, PA, US. IFN-g levels were calculated by reference to a standard curve constructed with recombinant cytokine (R&D Systems, Minneapolis, MN, US). Sensitivity of this method was 0.03 ng/ml. IL-4 was assayed also in a two-site ELISA, using rat anti-IL-4 mAb 11b11 (ATCC) and biotynilated mAb BVD6 (kind gift of Dr Phillip Scott, University of Pennsylvania). IL-4 levels were calculated by reference to a standard curve constructed with recombinant cytokine (R&D Systems). Sensitivity of this method was 0.06 ng/ml. All monoclonal antibodies were produced in our laboratory, from ascitis from nu/nu mice of various backgrounds. TNF production was estimated in a bioassay using WEHI cells as described previously (Lattime et al. 1988). Statistical analysis - All experiments used groups of 4 animals and were repeated twice with similar results. Results are represented as mean + SEM. Differences between groups were compared using the Student's t test. RESULTS Course of L. braziliensis infection in IL-12p40 -/- mice - Previous studies have demonstrated that IFN-g is essential to the control of L. braziliensis infection in BALB/c mice. Since IL-12 is involved in the induction of IFN-g production by NK as well as T cells, we decided to evaluate the role of IL-12 in the control of this parasite. While lesions in C57BL/6 mice inoculated with 1 x 107 stationary phase promastigotes begin to decrease in size 3 weeks after infection and are completely healed after 6 to 7 weeks, IFN-g -/- mice developed severe uncontrolled lesions and had to be sacrificed after 4 weeks of infection (Fig. 1). On the other hand, lesion growth in IL-12p40 -/- mice infected with L. braziliensis reached a maximum after 4 weeks of infection without further increase in size for at least another 8 weeks, suggesting a partial control of the parasite even in the absence of IL-12. Lesion development correlated with parasite load at the site of infection. As shown in Fig. 2, while in C57BL/6 mice a decrease in lesion parasitism from 3 to 12 weeks of infection was observed (3.7 ± 0.7 at 3 weeks and not detected at 12 weeks), the parasite load in lesions from IL-12p40 -/- slightly increased during the same time period (4.7 ± 0.4 at 3 weeks versus 5.7 ± 0.5 at 12 weeks p < 0.05). On the other hand, IFN-g -/- mice presented much higher numbers of parasites in their lesions (8.4 ± 0.9 at 4 weeks). Although parasite growth in IL-12p40 -/- mice appeared to be controlled at the site of inoculation, dissemination to other organs was not blocked in these animals. Histological examination of liver and spleen demonstrated the presence of the parasite in these organs. Furthermore, small metastatic skin lesions were observed in some animals later in the course of infection (data not shown). No dissemination of the parasite to viscera or presence of metastatic lesions was observed in infected C57BL/6 control mice. Similar results were obtained with animals inoculated with 1 x 105 promastigotes with regard to both lesion development and tissue parasitism. C57BL/6 mice developed small lesions that completely healed after seven weeks of infection. On the other hand, IL-12p40 -/- mice developed chronic lesions identical in size to those of mice infected with 1 x 107 parasites (data not shown). The parasite load of animals infected with 1 x 105 promastigotes was 1 x 102 and 1 x 107.5 for C57BL/6 and IL-12p40 -/- animals, respectively, at 12 weeks of infection. Cytokine production - In order to investigate the immune response to the parasite, lymph node cells were stimulated in vitro with L. braziliensis antigen and cytokine production measured after 24 or 72 h. As expected, stimulation of lymph node cells from C57BL/6 mice infected for 3 weeks induced the production of high levels of IFN-g (75 ± 39 ng/ml) while only very low levels of this cytokine were detected in the supernatants of IL-12p40 /- stimulated lymph node cells (0.8 ± 0.4 ng/ml) (Fig. 3a). At 12 weeks of infection a decrease in the production of IFN-g was detected in the lymph node of C57BL/6 mice (14.8 ng/ml), probably due to the decrease in the local response caused by the control of the parasite. The level of IFN-g production by cells IL-12p40 -/- mice at this point was similar to that detected at 3 weeks of infection (0.6 ± 0.3 ng/ml). With regard to IL-4 production, even though slightly higher levels were detected in supernatants from IL-12p40 -/- mice in comparison to C57BL/6 controls, especially at 3 weeks of infection, these differences were not statistically significant (Fig. 3b). Furthermore, IL-12p40 -/- mice produced less IL-4 at 12 weeks of infection than at 3 weeks (p < 0.05) indicating that, as opposed to what has been shown in L. major infected mice (Mattner et al.1996, 1997a), the lack of IL-12 during infection with L. braziliensis does not induce a strong polarization towards a Th2 type response. In addition, the levels of IL-4 detected in IL-12p40 -/- mice are much lower than those found in BALB/c mice infected with L. major (1-2 ng/ml, data not shown). An interesting observation was made when 24 h supernatants from infected mice were assayed for the presence of tumor necrosis factor (TNF). At 3 weeks after infection, levels of TNF detected in stimulated supernatants from both C57BL/6 and IL-12p40 -/- were 0.16 ± 0.04 and 0.34 ± 0.10 ng/ml, respectively. After 12 weeks of infection, however, a large increase in the levels of TNF was detected in supernatants from L. braziliensis-infected C57BL/6 mice (3.5 ± 1.5 ng/ml). This level of TNF production is greater than what is usually observed in L. major-infected C57BL/6 mice (Vieira et al. 1996). In IL-12p40 -/- infected with L. braziliensis however, this increase was not detected (0.8 ± 0.6 ng/ml) (Fig. 3c). When animals were infected with 1 x 105 promastigotes, a similar cytokine profile was observed in IL-12p40 -/- mice, i.e. very low levels of IFN-g, IL-4 and TNF, 12 weeks after infection. On the other hand, a decreased production of IFN-g and TNF by lymph node cells from C57BL/6 mice inoculated with 1 x 105 parasites was observed at 12 weeks of infection (1.2 and 0.02 ng/ml, respectively). No change in the production of IL-4 was observed. It is not clear however, whether this decreased production of inflammatory cytokines is responsible for the comparatively higher (but still very low) numbers of parasites found in C57BL/6 mice inoculated with 1 x 105 promastigotes. DISCUSSION The role of IL-12 in the induction of protection against infection by Leishmania has been extensively studied over the past 10 years. Several studies demonstrated that IL-12 production or its administration during the first weeks of infection by L. major is fundamental to the development of a protective response against this parasite (Heinzel et al. 1993, Sypek et al. 1993, Vieira et al. 1994, Scharton-Kersten & Scott 1995, Mattner et al. 1996), due to the importance of this cytokine in the induction of IFN-g production by NK cells and the generation of a Th1 response (Chan et al. 1991, 1992, Hsieh et al. 1993, Macatonia et al. 1993, Seder et al. 1993, Tripp et al. 1993). In addition, studies with IL-12 deficient mice infected with L. major demonstrated that this cytokine is important not only for the establishment, but also for the maintenance of the protective response against the parasite (Stobie et al. 2000, Park et al. 2000). However, recent studies have demonstrated that resistance to other intracellular pathogens can be achieved in the absence of IL-12 (Scharton-Kersten et al. 1996, Brombacher et al. 1999, Del Rio et al. 2001) and that this resistance may be mediated by IFN-g-producing CD4+ T cells, suggesting that IL-12 may be more important in the effector phase of the response rather than in its priming (Jankovic et al. 2002). Infection of several inbred mouse strain by L. braziliensis, the main causative agent of cutaneous leishmaniasis in Brazil, results in complete control of parasite growth (Childs et al. 1984, Lima et al. 1999). Contrary to what has been observed with L. major, infection by L. braziliensis does not induce a strong IL-4 production early in the course of the disease (DeKrey et al. 1998). In view of this observation, we decided to investigate how this inability to induce a strong Th2 response would affect the course of infection in the absence of a Th1 inducer, i.e. IL-12. The present study shows that infection of IL-12p40-/- mice with L. braziliensis results in development of chronic lesions that fail to heal but, nevertheless, do not develop progressively as demonstrated for mice infected with L. major (Mattner et al. 1996, 1997b), suggesting that in the absence of high levels of IL-4, mechanisms other than induction of Th1 cells by IL-12 may be relevant for parasite control. In view of the fact that IFN-g production is paramount to the control of this parasite (Fig. 1, DeKrey et al. 1998), we can assume that the partial control of the lesions in IL-12p40 -/- mice is dependent on the presence of some level of IFN-g production. We were able to detect low levels of IFN-g production by lymph node cells from infected IL-12p40 -/- animals. Although we did not determine the source of this IFN-g production, the experimental conditions used to stimulate the cells suggest that it may be associated to CD4+ T cells (Belkaid et al. 2002). It has been recently demonstrated that, although delayed, generation of Th1 cells in the absence of IL-12 is possible (Jankovic et al. 2002). It was suggested that IL-18 would be driving this differentiation (Okamura et al. 1995). The delayed generation of a small population of Th1 cells in IL-12p40 -/- mice infected with L. braziliensis would allow for the partial control of the lesion at a chronic stage of the disease. Alternatively, CD8+ T cells could be the effector cells controlling parasite growth in our model. Production of IFN-g by these cells has been reported to be independent of the action of IL-12 (Wan et al. 2001). The participation of CD8+ T cells in the control of Leishmania infection is well established, both in mice and humans (Coutinho et al. 1996, 1998, Huber et al. 1998, Holaday 2000, Belkaid et al. 2002). The involvement of CD4+ and/or CD8+ cells in the control of the infection by L. braziliensis in IL-12p40 -/- mice is currently being investigated. Lastly, our results show an interesting finding concerning TNF production by lymph node cells from infected mice (Fig. 3, bottom panel). Elevated levels of TNF were detected in cultures (both C57BL/6 and IL-12p40 -/-) as compared to those reported for mice infected with L. major under the same culture conditions (Vieira et al. 1996, Wei et al. 1999). Interestingly, patients with mucocutaneous leishmaniasis, a disease usually caused by L. braziliensis, show elevated levels of TNF in the serum (da Cruz et al. 1996). It would be interesting to verify the role of this cytokine in the control of infection in this model. ACKNOWLEDGMENTS To Maria Helena Alves de Oliveira, Antônio Vaz Mesquita, and Antônio Celso Torres for dedicated animal care. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc04052f1.jpg] [oc04052f2.jpg] [oc04052f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}