 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Vol. 99, Suppl. 1, August, 2004, pp. 37-43 Vaccination with Antioxidant Enzymes Confers Protective Immunity against Challenge Infection with Schistosoma mansoni Philip T LoVerde+, Claudia Carvalho-Queiroz, Rosemary Cook Department of Microbiology
and Immunology, School of Medicine and Biomedical Sciences, State University
of New York, Buffalo, NY 14214, US Financial support: NIAID grant AII8867 Received 28 May
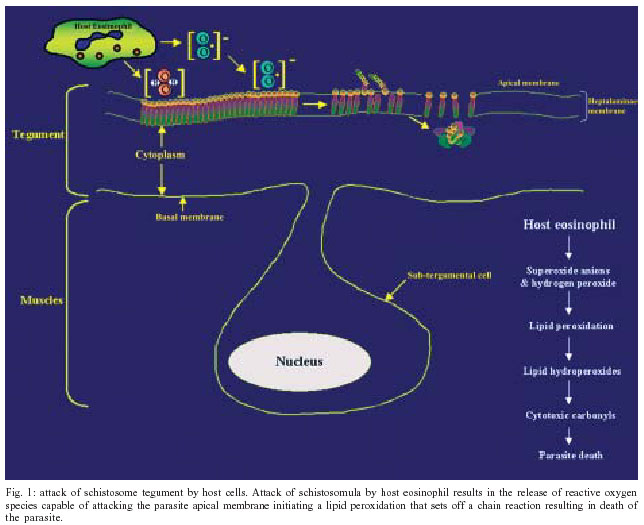
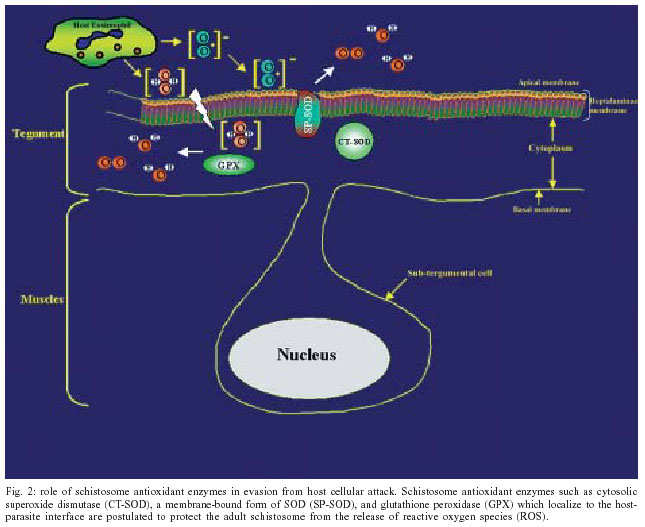
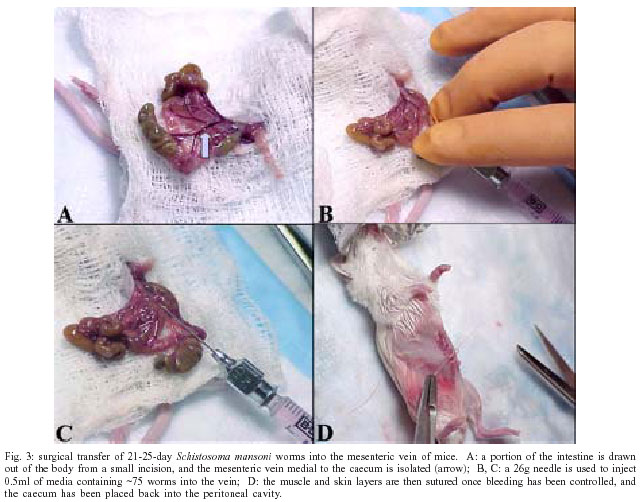
2004 Code Number: oc04084 Schistosoma mansoni, an intravascular parasite, lives in a hostile environment in close contact with host humoral and cellular cytotoxic factors. To establish itself in the host, the parasite has evolved a number of immune evasion mechanisms, such as antioxidant enzymes. Our laboratory has demonstrated that the expression of antioxidant enzymes is developmentally regulated, with the highest levels present in the adult worm, the stage least susceptible to immune elimination, and the lowest levels in the larval stages, the most susceptible to immune elimination. Vaccination of mice with naked DNA constructs containing Cu/Zn cytosolic superoxide dismutase (CT-SOD), signal-peptide containing SOD or glutathione peroxidase (GPX) showed significant levels of protection compared to a control group. We have further shown that vaccination with SmCT-SOD but not SmGPX results in elimination of adult worms. Anti-oxidant enzyme vaccine candidates offer an advance over existing vaccine strategies that all seem to target the larval developmental stages in that they target adult worms and thus may have therapeutic as well as prophylactic value. To eliminate the potential for cross-reactivity of SmCT-SOD with human superoxide dismutase, we identified parasite-specific epitope-containing peptides. Our results serve as a basis for developing a subunit vaccine against schistosomiasis. Key words: schistosomiasis - DNA vaccination - antioxidant - protection Schistosomiasis is a major cause of morbidity in the world, afflicting over 200 million people and over 600 million people are at risk of contracting the disease (Chitsulo et al. 2000, Bergquist 2002). Pathogenesis results from eggs produced by adult worm pairs that inhabit the portal circulation of humans. The severity of disease is a function of the worm burden (number of egglaying adult worms) and consequently, the inflammatory response to deposited eggs. The intensity of infection (worm burden) increases with exposure, as adult worms do not multiply within the human host. It turns out that approximately 10% of those infected exhibit severe forms of disease. Although drug treatment is available (Butterworth et al. 1992), often there are very high reinfection rates that make the treatment very costly especially in developing countries. With time, infection rates reach pre-treatment levels once drug treatment ceases (Gryseels et al. 1994) and the risk of drug resistance development is eminent. Therefore a vaccination program that would reduce the worm burden to prevent severe disease and reduce parasite transmission is desperately needed (Taylor 1991, Bergquist 1995). Although desirable, a sterilizing vaccine is not essential to attain these goals. The ultimate goal of our research is the development of an efficacious vaccine against schistosomiasis that contains parasite-specific epitopes. EVIDENCE THAT ANTIOXIDANTS PLAY A ROLE IN IMMUNE EVASION Although immunity in schistosomiasis has been studied extensively in experimental models and in human populations (Butterworth et al. 1992, Capron et al. 1992, Dessein et al. 1992, James 1992, Coulson 1997), immune correlates of resistance remain undefined. This is primarily because the various animal model systems and human studies have provided conflicting evidence regarding the immunological factors responsible for resistance. However, two paradigms have emerged from the various immune models of schistosomiasis studied to date. The first paradigm is that the larval stage (schistosomulum) is the most susceptible and the target of immune elimination (Smithers 1982, Smithers &Terry 1969, LoVerde 1998). Evidence indicates that larval parasite killing primarily involves a cellular response that includes monocytes, macrophages, eosinophils, and platelets (Fig. 1) that is potentiated by cytokines and/or antibodies (Butterworth 1984, Capron 1987, Coulson & Wilson 1997, LoVerde 1998). As part of the cell cytotoxic response against the schistosomula, it is thought that reactive oxygen species (ROS) such as superoxide anion and hydroxyl radicals are produced (see Fig. 1 and LoVerde 1998 for review). The second paradigm is that the adult parasite seems to be able to evade immune elimination through the evolution of several defense mechanisms (Smithers & Terry 1969, Smithers 1982, Maizels et al. 1993, Pearce & Sher 1987, LoVerde 1998). We postulate that in order to survive in the portal circulation of the liver, the adult worm stage has evolved a defense against the immune attack that involves the expression of antioxidant enzymes (Fig. 2), among other mechanisms (Callahan et al. 1988, Brophy & Pritchard 1992, Maizels et al. 1993, Mei and LoVerde 1997, LoVerde 1998). Antioxidant enzymes protect an organism from reactive oxygen species-derived damage (James 1994, LoVerde 1998). One of our strategies has been to identify mechanisms that the schistosome adult parasite employs to evade the immune responses and use this information to identify and evaluate candidate vaccines. In this regard, in vitro studies have demonstrated that schistosomula are sensitive (95% killed) to oxidative killing, whereas adult worms exhibit much greater resistance (2% killed) to oxidative killing (Mkoji et al. 1988). We hypothesized that adult worms protect themselves against oxidant damage by producing antioxidant enzymes (LoVerde 1998). To begin to test this hypothesis, we reasoned that the antioxidant enzymes should be developmentally regulated. We went on to demonstrate that expression of the schistosome antioxidant enzymes (Cu-Zn superoxide dismutase, SOD; glutathione S peroxidase, GPX) is developmentally regulated such that the lowest levels of gene expression (as measured by transcription) and enzyme specific activity were in the larval stages, the most susceptible to immune killing, and highest in adult worms, the least susceptible to immune elimination (Hong et al. 1992, Maizels et al. 1993, Mei & LoVerde 1995, 1997, LoVerde 1998). We next hypothesized that if antioxidant enzymes were to protect the adult worms from ROS, the antioxidant enzymes should localize at the host-parasite interface. Indeed, immunolocalization studies demonstrated that SmCT-SOD, SmSP-SOD, and SmGPX localized to the tegument (host-parasite interface) of adult but not in larval parasites supporting the notion that antioxidant enzymes are important in immune evasion by adult schistosome parasites (Mei & LoVerde 1997). Thus, antioxidant enzymes are hypothesized to play a role in protecting the parasite from damage caused by reactive oxygen species such as superoxide radical anion and hydroxyl radicals (Callahan et al. 1988, Brophy 1992, LoVerde 1998, Maizels et al. 1993, James 1994). As catalase activity has never been demonstrated in schistosomes (Mkoji et al. 1988a,b), we further hypothesize that the first line of enzymatic cellular defense against tegument attack (lipid peroxidation) in adult worms that results from the release of reactive oxygen species by host cells, involves parasite SOD and GPX (Fig. 2 and LoVerde 1998). ANTIOXIDANTS ARE VACCINE CANDIDATES To provide direct evidence that antioxidant enzymes were important in immune evasion and thus were viable candidate vaccines, we used DNA vaccination strategies to evaluate the efficacy of DNA constructs encoding either Cu/Zn cytosolic superoxide dismutase (SmCT-SOD), signal peptide-containing SOD (SmSP-SOD), or glutathione peroxidase (SmGPX) to be protective against S. mansoni infection in a murine challenge model of S. mansoni. Employing different doses of plasmid cDNA constructs, mice exhibited a significant level of worm burden reduction when challenged with S. mansoni cercariae after immunization with SmCT-SOD (54%) and SmGPX (43.4%) from six independent experiments (Shalaby et al. 2003). The WHO has identified several candidate vaccine antigens for independent evaluation, but none fulfilled the required standards in trials with mice. One of the criteria was a worm burden reduction of greater than 40% in a murine challenge model (Bergquist 1995). We have demonstrated that both SmCT-SOD and SmGPX are each capable as DNA-based (plasmid or vaccinia virus vehicles) vaccines to consistently induce significant levels of protection in an S. mansoni murine-challenge model, greater than the target 40% reduction in worm burden compared to controls set as a minimum by the WHO. ANTIOXIDANTS TARGET ADULT WORMS Once we demonstrated antioxidants could be vaccine candidates and that the stage with highest levels of expression and activity of those enzymes were the adult worms, we challenged the second paradigm that argues that adult worms are the least susceptible to immune killing. We hypothesized that adult worms could be a target of immune elimination following vaccination with antioxidants (Cook et al., in press). In order to test our hypothesis, after vaccination of BALB/c mice with our vaccine candidates, we used a surgical technique (Cioli 1976) to transfer exact numbers of adult worms aged 21-23 days directly into the mesenteric circulation (Fig. 3). Challenging the mice in this manner allowed us to evaluate the influence of our vaccination on liver-stage worms while circumventing all schis-tosomule stages and immune responses elicited by them (Cook et al., in press). In support of our hypothesis, we demonstrated that mice vaccinated with pcSmCT-SOD were significantly protected over controls (mean of 39%) in three independent experiments. Surprisingly, mice immunized with pcSmGPX did not experience significant decrease in worm burden, although previous studies shown significant protection in pcSmGPX vaccinated mice when challenged with cercariae (Shalaby et al. 2003). The reasons for the lack of protection observed against adult worms in pcSmGPX vaccinated mice, is currently under investigation. The average 39% protection observed in the three experiments using pcSmCT-SOD in the worm transfer model (when compared to the one using cercarial challenge) suggest that the majority of the parasite target by the immune responses elicited through vaccination with pcSmCT-SOD are worms 21 days and older. These data challenge the existing paradigms in schistosome immunology as well as offer an advance in schistosome vaccinology as immune attrition of adult worms could lead to the development of a therapeutic as well as prophylactic vaccine. POTENTIAL FOR NATURAL BOOSTING In order to determine what type of immune response(s) was elicited by DNA vaccination with pcSmCT-SOD and pcSmGPX, antibody responses to specific antigen were assessed in each experimental group and compared to the control groups. We found that there was a significant increase in specific total IgG responses in sera from mice vaccinated with pcSmCT-SOD compared to controls following challenge (Cook et al., unpublished observations). This implies that the specific antibody response primed by DNA vaccination in these two groups is boosted by infection. This "natural" boosting would be beneficial in endemic areas where individuals are continuously exposed to the parasite. To understand immune mechanisms stimulated in this animal model, various cytokine assays were performed at both the pre-challenge and pre-perfusion time points using spleen cells of mice in recall experiments from each experimental and control group. Based on our cytokine data, there was an overall decrease in intracellular Th1 cytokines from the pre-challenge to the pre-perfusion time point in all groups, and an increase in Th2 cytokines in pcXPG (control), pcSmCT-SOD, and pcSmGPX groups, consistent with previous reports on infection with schistosomes (Pearce et al. 1991, Vella & Pearce 1992, Wynn & Cheever 1995). When comparing cytokine production upon antigen stimulation in the unprotected groups (pcXPG and pcSmGPX) to the protected group (pcSmCT-SOD), two main trends were noted. First, the increased levels of both the IFN-g cytokine and IgG2a antibody observed in pcCT-SOD mice in this model may point to a possible role for a Th1/Tc1 response in protection, since interferon (IFN-g) stimulates the production of IgG2a. However, these findings do not rule out the possibility of other immune factors being involved in protection. The second trend noted was that the frequencies of lymphocytes producing Th2 cytokines such as interleukin (IIL-4), IL-5, and IL-10 increased in response to specific, egg, and worm antigen in the pcSmCT-SOD, when compared to the pcXPG and pcSmGPX groups following challenge. This evidence suggests that a mixed Th0 response, rather than a polarized response may be needed for adequate protection (Wynn & Hoffmann 2000). Another possibility is that while a polarized response is effective in adult worm killing, a different type of response is elicited to other worm or egg antigens. To better define the immune response(s) responsible for the protection observed in this model, mice with defined genetic deletions may prove a useful tool. Overall, the cytokine and antibody data support our hypothesis that ADCC (possibly using IgG2a to activate eosinophils) may play an important role in protection against a S. mansoni infection. VACCINATION WITH PARASITE CT-SOD INDUCES HOST CROSS-REACTIVE ANTIBODIES Autoimmune responses elicited by cross-reactive antibodies could lead to tissue damage such as seen in individuals that develop antibody to streptococcal M antigens (Vashishtha & Fischetti 1993). Because of the shared identity between S. mansoni and host anti-oxidant enzymes, the possibility of generating antibodies to human epitopes was addressed. Our results show that there is the potential for immunization with the whole SmCT-SOD molecule to induce antibodies that react with denatured human SOD (Carvalho-Queiroz et al. 2004). However there was no cross-reactivity between antibodies from SmCT-SOD immunized animals or humans naturally exposed to S. mansoni infection and the non-denatured human SOD. The recognition of denatured but not native forms of myosin and SmPGK following immunization has been also described (Lee et al. 1995, Vashishtha & Fischetti 1993). DENTIFICATION OF PARASITE-SPECIFIC EPITOPES To circumvent the concern of epitopes formed from denatured host proteins to induce an autoimmune response, we analyzed the immunogenicity of linear synthetic peptides derived from SmCT-SOD. Analyses of the reactivity of specific antibody isotypes against these peptides were aimed to identify relevant B cell epitopes that could be used in a subunit vaccine (Joshi et al. 2000, Tallima et al. 2003). Synthetic peptides have been widely used for the evaluation of both B and T cell epitopes responsible for (or that could lead to) protection against other pathogens, including Schistosoma (Reynolds et al. 1992, 1994, Eggleton et al. 2000, Joshi et al. 2000). We demonstrated that BALB/c mice vaccinated with SmCT-SOD developed high levels of antibodies directed mainly to the SmCT-SOD derived peptide 3 (Pep 3), which included amino acid residues 64-83 (Carvalho-Queiroz et al. 2004). Since the efficacy of a given vaccine depends on proper antigen processing and presentation within the host cells as well as on the need and/or type of vaccine adjuvant required, as reviewed in O'Hagan et al. (2001a,b), we also evaluated the reactivity of antibody isotypes from mice immunized by different formulations of SmCT-SOD. The biodegradable microspheres such as polylactide-co-glycolides (PLG); poly-D, L-lactide-co-glycolic (PLGA) and polylactic acid (PLA) have the ability to control the rate of release of encapsulated antigens (O'Hagan 1998) thus enhancing their adjuvant effect (O'Hagan et al. 1991a,b). Their properties have been explored successfully from cancer therapy (Egilmez et al. 1998, 2000), to infectious diseases (Okada & Toguchi 1995, Putney & Burke 1998, Rosas et al. 2002, Shi et al. 2002). Our results showed that although both adjuvant formulations used for delivery of SmCT-SOD proteins (encapsulated in PLA microspheres or absorbed in alum) elicited overall high levels of IgG antibodies, the mice vaccinated with SmCT-SOD encapsulated PLA microspheres developed higher levels of IgG2a anti-SmCT-SOD and anti-peptide 3 (Pep 3) antibodies when compared to alum adjuvant (Carvalho-Queiroz et al. 2004). The significance of the administration of SmCT-SOD with different adjuvant formulations (including DNA, protein and peptides) as well as the role of specific antibodies or cellular responses and the development of protection against challenge with S. mansoni are unknown, and currently under investigation. We have been able to provide evidence that one mechanism adult worms use to evade the immune response is the production of antioxidant enzymes at the host-parasite interface. Schistosome antioxidant enzymes are excellent vaccine candidates. SmCT-SOD and SmGPX, each consistently confer greater than 40% protection, the WHO minimum requirement, when delivered as a DNA-based vaccine. Furthermore, our studies show that adult worms are subject to immune elimination by vaccination with SmCT-SOD. The advance for the field of schistosome vaccinology is that SmCT-SOD has the potential to act as a therapeutic vaccine that eliminates established worm infections as well as offer protection as a prophylactic vaccine. The potential adverse effect of SmCT-SOD inducing host cross-reactive immune responses was demonstrated. To overcome this disadvantage, parasite-specific epitopes were identified. The potential exists to develop a safe and efficacious subunit vaccine that can target established adult worms.
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc04084f3a-d.jpg] [oc04084f2.jpg] [oc04084f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}