 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 5, August, 2004, pp. 489-492 Production of L-asparaginase by Filamentous Fungi Maria Inez de Moura Sarquis/+, Edna Maria Morais Oliveira*, Alberdan Silva Santos**, Gisela Lara da Costa Departamento de
Micologia, Laboratório de Coleção de Fungos, Instituto
Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil
*Laboratório de Detecção de OGM, Embrapa Agroindústria
de Alimentos, Guaratiba, Rio de Janeiro, RJ, Brasil **Departamento de Engenharia
Química, Universidade Federal do Pará, Belém, PA, Brasil Received 11 November
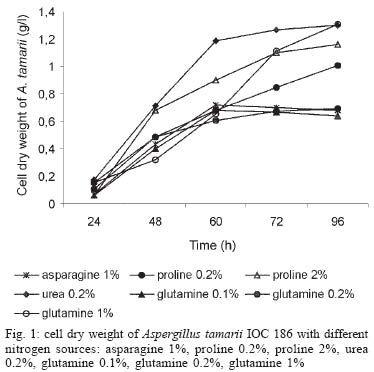
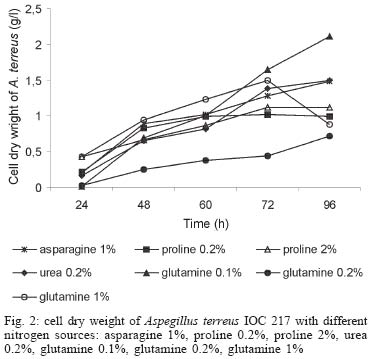
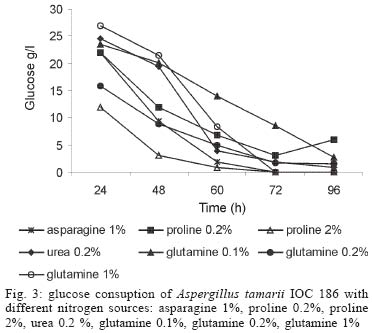
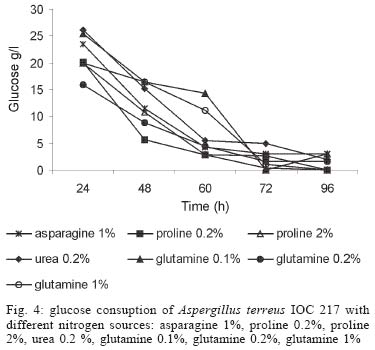
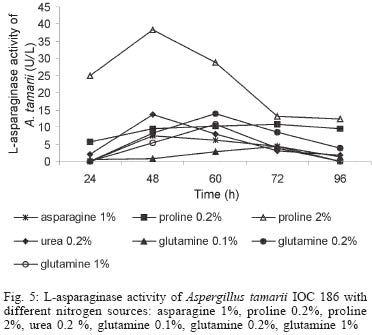
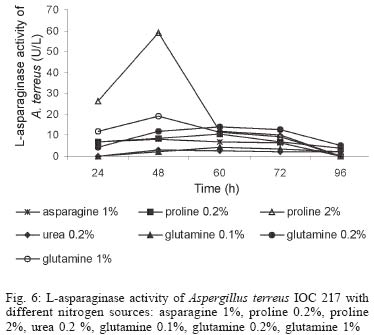
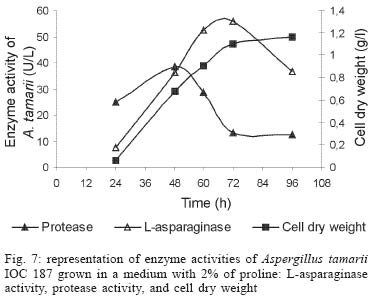
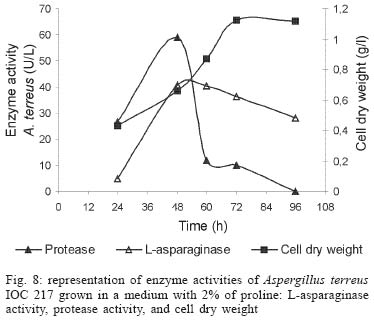
2003 Code Number: oc04105 L-asparaginase production was investigated in the filamentous fungi Aspergillus tamarii and Aspergillus terreus. The fungi were cultivated in medium containing different nitrogen sources. A. terreus showed the highest L-asparaginase (activity) production level (58 U/L) when cultivated in a 2% proline medium. Both fungi presented the lowest level of L-asparaginase production in the presence of glutamine and urea as nitrogen sources. These results suggest that L-asparaginase production by of filamentous fungi is under nitrogen regulation. Key words: filamentous fungi - production of L-asparaginase - Aspergillus tamarii - Aspergillus terreus Asparaginase (L-asparagine amido hydrolase, E.C. 3.5.1.1) is an anti-neoplastic agent, used in the lymphoblastic leukaemia chemotherapy. Neoplastic cells cannot synthetase L-asparagine due the absence of L-asparagine synthetase (Keating et al. 1993). For this reason the commonest therapeutic practice is to inject intravenously free enzyme in order to decrease the blood concentration of L-asparagine affecting selectively the neoplastic cells (Mitchell et al. 1994). However, L-asparaginase from bacterial origin can cause hypersensitivity in the long-term used, leading to allergic reactions and anaphylaxis (Reynolds & Taylor 1993). The search for other asparaginase sources, like eucaryotic microorganisms, can lead to an enzyme with less adverse effects. The importance of microorganisms as L-asparaginase sources has been focused since the time it was obtained from Escherichia coli and its antineoplastic activity demonstrated in guinea pig serum (Broome 1961, Mashburn & Wriston 1964, Roberts et al. 1966, Schawartz et al. 1966, Boyse et al. 1967). The E. coli and Erwinia enzymes were isolated, purified, and experimentally used as an anti-leukaemic agent in human patients (Clavell et al. 1986, Story et al. 1993). It de-monstrated high potential against children's acute lymphoblastic leukaemia (Hill et al. 1967, Oettgen et al. 1967). Several research groups have studied asparaginase production and purification in attempt to minimize impurities that produce allergenic reations (Campbell et al. 1967, Boss 1997, Gallagher et al. 1999). It has been observed that eukaryote microorganisms like yeast and filamentous fungi have a potential for asparaginase production (Wade et al. 1971, Wiame et al.1985, Pinheiro et al. 2001). For example, the mitosporic fungi genera such as Aspergillus, Penicillium, and Fusarium, are commonly reported in scientific literature to produce asparaginase (De-Angeli et al. 1970, Arima et al. 1972, Imada et al. 1973, Nakahama et al. 1973, Curran et al. 1985). L-asparaginase belongs to an amidase group that produces aspartic acid and ammonia by asparagine hydrolysis (Wriston & Yellin 1973, Capizzi et al. 1984). Here we present a simple and rapid assay method for the detection of asparaginase from different strains of Aspergillum, Penicillium, and Fusarium genera of fungi using the Gulati et al. (1997) method. After an initial screening, strains presenting high asparaginase activity were cultivated in submerged fermentation in order to determine the production of this enzyme in the presence of different nitrogen sources. Moreover the question whether asparaginase from filamentous fungi is nitrogen regulated as the enzyme produced by Saccharomyces cerevisae was also investigated (Oliveira et al. 2003). MATERIALS AND METHODS Chemical - Chemicals were of analytical grade and purchased from Difco (Michigan US). Analytical determinations were carried out at Laboratório de Coleção de Culturas de Fungos do Instituto Oswaldo Cruz and Laboratório de Tecnologia Enzimática do Instituto de Química, Universidade Federal do Rio de Janeiro, Brasil. Selection of filamentous fungi - The genera of Aspergillus, Penicillium, and Fusarium were previously selected by literature reports and 26 fungal species were used in this study. All of them were obtained from our departamental stock culture collection. The tested species had been preserved between 6 and 35 years in mineral oil. The spore and mycelial fragments of strains were cultivated in modified Czapek Dox medium as used for fungi (Saxena & Sinha 1981) which contained (gl-1 of distilled water): glucose, 2.0; L-asparagine,10.0; K2PO4 1.52; KCl, 0.52; MgSO4..7H2O 0.52; CuNO3. 3H2O, trace; ZnSO4. 7 H2O, trace FeSO4.7H2O,trace; agar 18.0, pH 6.2. Assay for screening of L-asparaginase - The methodology was based on Gulati et al. (1997) with the incorporation of phenol red in a stock solution prepared in ethanol with L-asparagine incorporated in the medium for the selection of the microorganism with the ability to produce L-asparaginase. Production of conidial suspension in submerged fermentation - A. tamarii and A. terreus strain were cultivated in Czapek agar plates at 26oC for 7 days. From these cultures, conidial suspensions were prepared with concentrations of 107 to 108 conidia ml-1. These suspensions were transferred to Erlenmeyer flasks with 200 ml of liquid Czapek-Dox modified with different nitrogen sources such as L-asparagine 1%, proline 0.2% or 2%, urea 0.2% and glutamine 0.1%, 0.2% or glutamine 1% (Merck) and submitted to an orbital shaker at 160 revolution min-1, at 30oC for 96 h. Determination of biomass production - Each culture was filtered each 24 h, on Whatman 2 paper filter. The growth curve was plotted with the cell dry weight versus time. The cell pellets were brought to 150oC for drying. Glucose residual determination - The supernatant from the culture was submitted to the Glucose-Oxidase-Peroxidase method (Beckman Glucose Analyzer, Kadish, 1968) for glucose determination. This analysis was carried out each 24 h, in order to monitor the glucose consumption in relation to biomass. Determination of L-asparaginase activity - This was performed by quantifying ammonia formation in a spectrophotometric analysis at 340 nm, using a Boering Mannhein Kit (Biopharm E1112732). Enzyme activities were determined in the culture filtrates at regular intervals of 24 h up to 96 h. Determination of protease activity - A volume of 0.5 ml of 10% azocasein were added in 0.5 ml of each 24 h culture filtrate was incubated at 30oC/30 min; 0.5 ml of 10% TCA was then added and the culture stored at -20oC for 5 min. The suspension was centrifuged at 3000 rpm for 10 min. The supernatant was analyzed by UV-vis spectrophotometer at 428 nm after addition of 0.5 ml of KOH 5N. The reaction blank was carried out by enzyme denaturing through incubating culture filtrate with 10% TCA for 5 min. After this time, the substrate 10% azocasein was added. RESULTS AND DISCUSSION This screening of filamentous fungi is based on the semi-qualitative method described by Gulati et al. (1997). Twenty-six strains belonging to Aspergillus, Penicillium, and Fusarium genera were submitted to this systematic investigation. In our screening the optimal chromogenic substrate concentration was 0.04%(V/V). This preliminary selection showed that only strains from Aspergillus genera presented asparaginase production. We select A. tamarii (strain IOC 186) and A. terreus (strain IOC 217) for further study. These 2 strains had been preserved during 23 years, showing that the viability and genetic stability remained unaltered. The L-asparaginase activity of A. terreus has been described in the literature (De-Angeli et al. 1970, Gulati et al. 1997, Zurita et al. 1971). The experiments were carried out as described as a kinetic study of biomass and L-asparaginase production using different nitrogen sources. Biomass production of A. tamarii with different nitrogen sources is shown at Fig. 1. The highest amount of biomass was obtained with urea 0.2% and glutamine 1% as nitrogen source, with proline 2% presenting the second best nitrogen source for growth. An unusual growth was obtained with glutamine that gave the same final biomass as urea, but the curve hyperbolic in contrast with that observed with urea that presented a parabolic. The biomass production of A. terreus with the different nitrogen sources presented different growth curves when compared with A. tamarii. In this case, the biomass production of A. terreus presented an averaged of 1.5 times greater than A. tamarii and glutamine 0.1% medium revealed itself as a good nitrogen source for the A. terreus growth (Fig. 2). The glucose consumption profiles were presented at Figs 3, 4. It was observed that glucose was totally consumed after 72 h for both strains. A. terreus IOC 217 strain in 2% proline medium showed the highest enzyme activity at 48 h, with 58.8 U/L whereas A. tamarii presented 38 U/L at the same conditions (Figs 5, 6). The determination of protease production was performed to explain the decrease of L-asparaginase activity. After 60 h, it was demonstrated that the protease activity could be responsible for decrease in L-asparaginase activity since protease levels increase when L-asparaginase levels decrease (Figs 7, 8). The results suggest that L-asparaginase production is nitrogen regulated, similar by to enzyme production in Saccharomyces cerevisae. As a rule, different nitrogen sources can be used to promote high enzyme production. Different amino acids were used since microorganisms can utilize a wide variety of nitrogen sources. The nitrogen regulation mechanism in filamentous fungi has been carefully studied. It has been observed that the structural gene expression is under GATA factor control as the activators ARE-A and NIT-2, regulate gene transcription according to nitrogen availability (Caddick et al. 1994). The proline 2% medium was observed to be the best condition for L- asparaginase production (Figs 5, 6). Indeed, the highest L-asparaginase activity level was present by A. terreus at 48 h. However, these activity levels could be improved with the utilization of protease inhibitors. ACKNOWLEDGEMENTS To Dr Elba Pinto da Silva Bon from the Universidade Fede-ral do Rio de Janeiro for making available the equipment for this analyse. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc04105f7.jpg] [oc04105f3.jpg] [oc04105f2.jpg] [oc04105f1.jpg] [oc04105f8.jpg] [oc04105f4.jpg] [oc04105f5.jpg] [oc04105f6.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}