 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 5, August, 2004, pp. 525-530 Antileishmanial IgG and IgE Antibodies Recognize Predominantly Carbohydrate Epitopes of Glycosylated Antigens in Visceral Leishmaniasis AM Atta/+, R Colossi/++, MLB Sousa-Atta, SMB Jeronimo*, MDSB Nascimento**, GF Bezerra**, G Orge***, EM Carvalho*** Departamento de
Análises Clínicas e Toxicológicas, Laboratório
de Pesquisa em Imunologia, Faculdade de Farmácia, Universidade Federal
da Bahia, Rua Barão de Geremoabo s/no, Campus de
Ondina, 40171-970 Salvador, BA Brasil *Departamento de Bioquímica,
Universidade Federal do Rio Grande do Norte, Natal, RN, Brasil **Departamento
de Patologia, Universidade Federal do Maranhão, São Luiz, MA,
Brasil ***Serviço de Imunologia do Hospital Universitário Professor
Edgard Santos, Universidade Federal da Bahia, Salvador, BA, Brasil Received 12 January
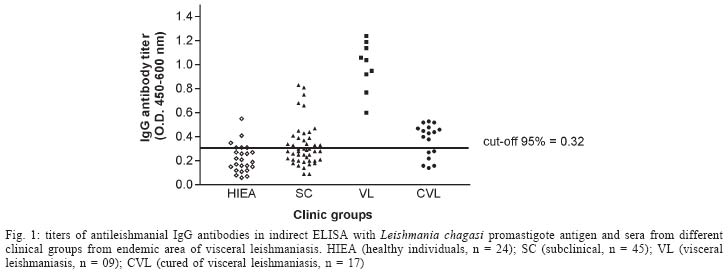
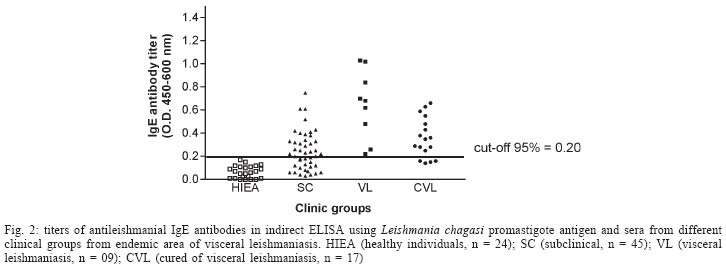
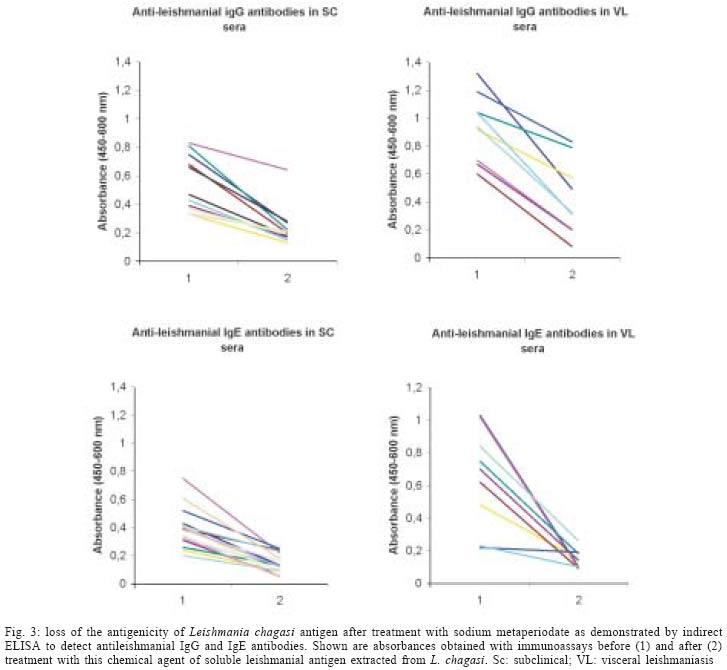
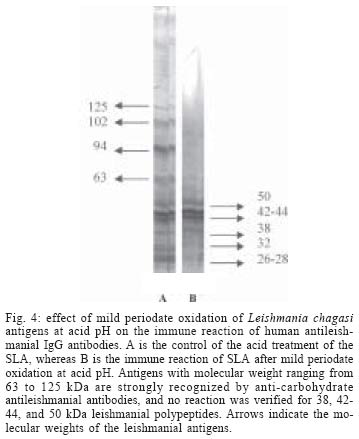
2004 Code Number: oc04112 The specificity of human antileishmanial IgG and IgE antibodies to glycosylated antigens of Leishmania chagasi was evaluated. An ELISA was performed with soluble leishmanial antigen (SLA) and a panel of 95 sera including samples from patients with subclinical infection (SC) and visceral leishmaniasis (VL), subjects cured of visceral leishmaniasis (CVL), and from healthy individuals from endemic areas (HIEA). Antileishmanial IgG were verified for 18 (40%) of 45 SC subjects (mean absorbance of 0.49 ± 0.17). All nine sera from VL patients had such antibody (0.99 ± 0.21), while 11 (65%) of 17 CVL individuals were seropositive (0.46 ± 0.05). Only three (12%) of 24 HIEA controls reacted in IgG-ELISA. Antileishmanial IgE was detected in 26 (58%) of 45 SC patients (0.35 ± 0.14), and in all VL patients (0.65 ± 0.29). These antibodies were also detected in 13(76%) of 17 CVL subjects (0.42 ± 0.14) while all HIEA controls were seronegative. There was no correlation between antileishmanial IgG and IgE antibody absorbances. Mild periodate oxidation at acid pH of SLA carbohydrates drastically diminished its antigenicity in both IgG and IgE-ELISA, affecting mainly the antigens of 125, 102, 94, and 63 kDa as demonstrated by western immunoblotting. Key words: visceral leishmaniasis - anti-carbohydrate antibodies - IgG - IgE Leishmania chagasi is an obligatory intracellular parasite that causes human visceral infection in South and Central Americas. The human natural habitat of this protozoan is the mononuclear phagocyte from bone marrow, spleen, lymph nodes, gut, and liver. Exuberant splenomegaly, hepatomegaly, blood pancytopenia, polyclonal activation, and hypergammaglobulinemia characterize visceral leishmaniasis (VL). A high mortality is observed in VL patients when antimonial chemotherapy is omitted or ineffective (Badaró & Duarte 1997). In addition to the classical clinical form of the disease, studies in areas of L. chagasi transmission have identified individuals that were exposed to Leishmania and presented anti-Leishmania IgG antibodies or a positive intradermal Montenegro reaction (Badaró & Reed 2001). Since these individuals had no clinical manifestation typical of VL they were considered having a subclinical L. chagasi infection (SC). An intense polyclonal antibody synthesis against Leishmania polysaccharides, glycolipids, glycoproteins, and also proteins is easily detected in VL (Badaró & Reed 2001). Such antibodies are well demonstrated in patient sera by immunoassays like direct agglutination test (DAT), indirect fluorescent antibody test (IFAT), enzyme-linked immunosorbent assay (ELISA) or western immunoblot-ting, having different isotypes and affinities, which may be associated with clinic and immune features of the Leishmania infection (Ghosh et al. 1995, Shiddo et al. 1996, Anam et al. 1999). Previously, others and we reported the occurrence of antileishmanial IgE antibodies in sera from VL patients, which were associated with active disease (Atta et al. 1998, Anam et al. 1999). However, a study has demonstrated that specific IgE antibodies to cross-reactive carbohydrate determinants (CCDs) shared by glycosylated antigens from different sources affect the in vitro diagnosis of allergic diseases (Mari et al. 1999). Also, the specific reactivities of IgE antibodies with glycoprotein antigens are improved after mild periodate oxidation at acid pH of their glycosylated epitopes, which destroys carbohydrate residues and does not affect the antigenicity of peptide epitopes (Afferni et al. 1999, Mari et al. 1999). Although several reports already demonstrated the antigenicity of Leishmania glycolipids and glycoproteins during the IgG immune response observed in leishmaniasis (Kutner et al. 1991, Palatnik de Sousa et al. 1993, Shreffler et al. 1993), the reaction of glycosylated epitopes with antileishmanial IgE antibodies in this disease is unknown. The aim of this work was to investigate the occurrence of IgG and IgE antibodies to glycosylated epitopes of L. chagasi pro-mastigote antigens in the antileishmanial immune response of subjects presenting the subclinical and visceral forms of leishmaniasis. MATERIALS AND METHODS Sera - The serologic survey of antileishmanial IgG and IgE antibodies was carried out with a panel of 95 human sera from Brazilian endemic areas of L. chagasi (Rio Grande do Norte and Maranhão), including serum samples from 24 healthy individuals (HIEA), from 45 subjects having SC infection, 9 sera from patients suffering of VL and sera from 17 persons cured of VL by antimonial therapy (CVL). Subclinical L. chagasi infection was defined by clinical exams, the presence of serum IgG antibodies to Leishmania and/or a positive Montenegro intradermal test for leishmaniasis in the absence of clinical diagnosis or previous history of VL. L. chagasi antigen - In order to obtain leishmanial soluble antigen (SLA), 109 stationary promastigotes from a cloned human strain of L. chagasi (MHOM-BR 86-BA), grown in LIT medium supplemented with calf fetal serum and antibiotics, were washed three times in PBS (10 mM phosphate buffer containing 150 mM NaCl, pH 7.4) and disrupted with 6 mM CHAPS (3-[(3-Cholamidopropyl) dimethylammonio]-1-propane-sulfonate) in 50 mM Tris-HCl buffer containing 150 mM NaCl, pH 7.5, and protease inhibitors (PMSF, leupeptin, antipain, ortho-phenantroline, and TPCK). After centrifugation at 13,000 g for 15 min at 4°C, the supernatant was evaluated for protein content and stored at - 20°C until used as SLA. IgG and IgE immunoassays - Antileishmanial IgG and IgE antibodies were determined through immunoassays performed into round-bottom wells of polystyrene microtiter plates (Nunc, Maxsorp, Denmark) coated with 500 ng of L. chagasi protein. After antigen sensitization, the free reactive sites of the wells were blocked with 1% BSA. Immune reactions of antileishmanial IgG antibodies were carried out with 100 µl of sera diluted at 1/500 in PBS-BSA for 1 h at room temperature (RT). After, the wells were thrice washed with PBS containing 0.05% Tween 20 (PBS-T) and incubated in the same conditions with 100 µl of goat anti-human IgG peroxidase conjugate (Sigma Chemical Co., St. Louis, MO) diluted 1/5000 in PBS-BSA. The wells were again washed three times with PBS-T, and the reactions were revealed with 100 µl of citrate-phosphate buffer containing hydrogen peroxide plus TMB (3,3', 5,5'-tetramethylbenzidine), during 30 min in the dark (RT). Following the addition of 50 µl of 2N sulfuric acid, the absorbances were measured at 450-600 nm in a Diamedix BP-12 ELISA reader. Antileishmanial IgE antibodies were investigated in IgG depleted sera by treatment with purified sheep IgG anti-human IgG (RF-Absorbentä, Dade Behring, Germany) as previously described (Sousa-Atta et al. 1999). To perform these assays, the wells coated with L. chagasi antigens were first incubated with 100 µl of IgG depleted sera diluted 1/6 in PBS-BSA for 18 h at 4°C. After the wells were washed with PBS-T, and incubated with 100 µl of goat anti-human IgE peroxidase conjugate (Sigma Chemical Co.) diluted 1/2500 in PBS-BSA for 2 h at 37o C. The reactions were revealed with hydrogen peroxide plus TMB as described in IgG immunoassay. The cut-off of both immunoassays were determined statistically (Frey et al. 1998) using 10 sera from healthy individuals (HI) from a free area of leishmaniasis (Salvador, Northeast Brazil). All of them were seronegative for rheumatic diseases (rheumatoid arthritis, SLE) and infections (American trypanosomiasis, syphilis, and viral hepatitis). The participation of glycosylated epitopes of L. chagasi antigens in the immune reactions with antileish-manial IgG and IgE antibodies was evaluated after mild periodate oxidation at acid pH of SLA (Woodward et al. 1985). In this test, leishmanial antigens were first adsorbed into the wells of ELISA plates and their carbohydrates oxidized to aldehyde with 100 µl of 10 mM sodium metaperiodate in 50 mM acetate buffer, pH 4.5, during 1 h at room temperature in the dark. After incubation, the wells were washed one time with sodium acetate buffer, being the new formed aldehyde groups reduced to alcohol through the incubation with 100 µl of 50mM sodium borohydride in PBS (Woodward et al. 1985). The wells were washed three times with PBS and immediately used in immunoassays. In order to evaluate the effect of acid treatment on the protein epitopes of leishmanial antigens, control wells coated with SLA were incubated with the pH 4.5 acetate buffer only, for 1 h. The proportion of carbohydrate contribution to the antibody immune reaction in ELISA was calculated comparing the absorbances obtained in the controls (100%) and tests. These analysis of carbohydrate antigenicity were carried out using 12 and 16 subclinical sera that were reactive in the immunoassays to detect antileishmanial IgG and IgE antibodies respectively, while all 9 VL serum samples were used in both tests. Western immunoblotting - In order to identify partially the leishmanial glycoproteins reacting with anti-carbohydrate antibodies, SLA was fractionated by 10% SDS-PAGE after treatment with 2-mercaptoethanol and electrotransferred to a PVDF membrane. After transfer, strips of the membrane containing the leishmanial antigens was first incubated with 0.5% Tween 20 in PBS to block residual free reactive sites (1 h at RT), thrice washed with PBS-T and then treated for mild periodate oxidation at acid pH as described for ELISA. The effect of acid treatment on the antigenicity of SLA was also investigated, treating strips of PVDF membrane containing fractionated SLA for 1 h at RT with the pH 4.5 acetate buffer only (Woodward et al. 1985). The membrane strips, control and tests, were washed with PBS-T, and incubated with VL sera diluted at 1/100 in PBS-T for 3h at 37oC. After, they were washed with PBS-T, and then incubated with goat anti-human IgG peroxidase conjugate (Sigma Chemical Co.) diluted at 1/2000 in PBS-T for 1 h at room temperature. The electroblots were revealed using as substrate for the HRP reaction diaminobenzidine (DAB) in PBS containing 0.03% hydrogen peroxide. Statistical analysis - The absorbances obtained in ELISA for antileishmanial IgG and IgE antibodies were reported as means ± SD and 95% Confidence Interval. Data were analyzed statistically by the t Student test, Mann-Whitney ranks sum test and also the Kruskal-Wallis one. Correlation analysis was determined by Spearman rank-order test. The level of significance set at P < 0.05. RESULTS Antileishmanial IgG antibodies were detected in 18 out of 45 (40%) sera from SC subjects, presenting this clinical group a mean absorbance of 0.49 ± 0.17 (CI 95% = 0.40 to 0.57). While all nine (100%) VL patients presented IgG antibodies to Leishmania (OD = 0.99 ± 0.21, CI 95% = 0.83 to 1.15), 11 out of 17 (65%) CVL subjects were seropositive in this test (OD = 0.46 ± 0.05, CI 95% = 0.43 to 0.49). Only three out of 24 (12%) healthy individuals from endemic areas were seropositive for antileishmanial IgG antibodies, showing ELISA absorbances ranging from 0.38 to 0.53 (Fig. 1). IgE antibodies to L. chagasi were detected in 26 out of 45 (58%) SC individuals, with a mean absorbance of 0.35 ± 0.14 (CI 95% = 0.30 to 0.41). In this group, 15 patients also had antileishmanial IgG antibodies, but 11 (24%) and three (7%) of them presented only IgE or IgG immune response against the parasite, respectively. As verified for antileishmanial IgG antibodies, all nine (100%) VL patients were also seropositive for IgE antibodies to Leishmania (OD = 0.65 ± 0.29, CI 95% = 0.42 to 0.87) while 13 out of 17 (76%) individuals cured of VL presented IgE antibodies against this parasite (OD = 0.42 ± 0.14, CI 95% = 0.34 to 0.51). IgE antibodies to Leishmania were not observed in the healthy inhabitants from endemic area (Fig. 2). The mean absorbances of antileishmanial IgG and IgE antibodies differed in SC, VL, and CVL groups (Kruskal -Wallis test, P < 0.0001 and P = 0.014, respectively), being both more elevated in the group of visceral leishmaniasis. There was no correlation in the Spearman rank-order test between the absorbances of the tests to detect antileish-manial IgG and IgE antibodies in SC group (r = 0.256, P > 0.05) or VL one (r = - 0.033, P > 0.05). While the antileishmanial IgG test using sera from VL and HIEA groups presented 100% sensitivity and 89% specificity, IgE immunoassay was 100% specific and sensitive with the same sera. Treatment of L. chagasi antigen by sodium meta-periodate caused a significant change in the absorbances of the immunoassays for antileishmanial IgG and IgE antibodies in SC and VL sera (Fig. 3). A mean absorbance of 0.24 ± 0.13 was verified in the ELISA for antileishmanial IgG antibodies in the SC group after antigen periodate oxidation while of 0.42 ± 0.27 was observed for VL sera. The controls of these reactions using antigen without periodate treatment presented means of antibody absorbances of 0.53 ± 0.20 and 0.94 ± 0.24 (Mann-Whitney test, P = 0.0002 and P < 0.0001, respectively). The absorbances of the ELISA for antileishmanial IgE antibodies also diminished after periodate oxidation of SLA, presenting the SC group a mean absorbance of 0.14 ± 0.07 (reaction control = 0.39 ± 0.14; Mann-Whitney test, P < 0.0001), while the VL one showed a mean of 0.14 ± 0.06 against 0.65 ± 0.29 of the reaction control (Mann-Whitney test, P < 0.0001). Such chemical treatment did not change the absorbances of the negative sera used to obtain the cut-off of both immunoassays. Western immunoblotting analysis (Fig. 4) showed that antileishmanial IgG antibodies reacted strongly with glycosylated epitopes of leishmanial polypeptides of 125, 102, 94, and 63 kDa, which lost their immunoreactivity after periodate oxidation of their carbohydrate residues. A discrete loss of antigenicity was also verified for 32 and 26-28 kDa antigens. The immune reactions of 55, 42-44, and 38 kDa leishmanial polypeptides with these antibodies were not affected by carbohydrate oxidation. On the other hand, there was no effect of acid treatment on the antigenicity of protein epitopes of SLA. While a high concentration of serum antileishmanial antibodies is observed in VL patients, individuals with the SC form present low titers of these immunoglobulins (Badaró & Duarte 1997). In this work, we demonstrated that SC individuals might produce simultaneously antileishmanial IgG and IgE antibodies during L chagasi infection. Furthermore most of the antibodies against L. chagasi recognized predominantly glycosylated epitopes. Previously others and we observed that both anti-leishmanial IgG and IgE antibodies could be detected simultaneously in VL patients (Atta et al. 1998, Anam et al. 1999), the titers of these antibodies decreasing after cure. Such decrease of antibody titers was also documented in a following-up study of Indian VL patients (Kumar et al. 2002). Although antileishmanial IgG antibodies were detected in some healthy individuals from the endemic areas of leishmaniasis, these antibodies may represent immune cross-reaction or document a naturally resolving subclinical leishmanial infection. One important observation here obtained was the intense reaction of antileishmanial IgG and IgE antibodies with glycosylated epitopes of Leishmania antigens, that presented a significant decrease in antigenicity after periodate oxidation of the carbohydrate residues. The binding of human IgE antibodies to carbohydrate epitopes of plant and invertebrate N-glycans is usually associated with the recognition of an alpha (1,3)-fucose linked to the proximal N-acetylglucosamine and also by a beta (1,2)-xylose linked to the core manose, being these antibodies induced after pollen sensitization and insect stings (Fotisch K & Vieths 2001, Petersen & Mundt 2001, van Ree 2002). Leishmania is a highly glycosylated parasite that has glycoconjugates that react with lectins presenting different carbohydrate specificities (Andrade & Saraiva 1999). Such exuberant glycosylation could contribute for crossed reactions between antileishmanial IgE antibodies and food and indoor allergens that shared the carbohydrate structures of these Leishmania glycocon-jugates. However, we verified in this study that IgE antileishmanial antibodies reacting strongly and specifically with glycosylated epitopes of L. chagasi promas-tigote are not detected in sera from healthy individuals from both leishmaniasis endemic and non-endemic areas. We already demonstrated that subjects presenting respiratory allergy do not react in antileishmanial IgE immunoassays (Atta et al. 1998). Additionally, we also verified that glycosylated epitopes of horseradish peroxidase bearing alpha (1,3)-fucose and beta (1,2)-xylose are not recognized by antileishmanial IgE antibodies in immunoassays using this enzyme as antigen (data not shown). Such exclusive antigenicity of glycosylated epitopes of L. chagasi glycoconjugates observed in human visceral leishmaniasis are not found in others parasite diseases such as human schistosomiasis, where glycosylated epitopes of glycoproteins of Schistosoma egg provoke significant crossed immune reactions in the serologic tests, which only are eliminated after antigen oxidation with sodium meta-periodate (Noya et al. 2000). Western immunoblotting of periodate oxidized SLA with antileishmanial IgG antibodies, showed that carbohydrate epitopes of glycoproteins with molecular weights of 63, 94, 102, and 125 kDa are strongly recognized in these immune reactions. On the other hand, the mild periodate oxidation of SLA at acid pH did not have any deleterious effect on protein epitopes of leishmanial antigens, which preserved their antigenicity after 1h incubation in pH 4.5 acetate buffer. Leishmanial antigens of 50, 42-44, and 38 kDa did not react with anti-carbohydrate antibodies, and probably contribute for the immune reactivity observed in the immunoassays for IgG and IgE antileishmanial antibodies after periodate oxidation of SLA. Some of these glycoproteins reacting with antileish-manial antibodies to glycosylated epitopes may be important leishmanial bioactive molecules, such as the surface metallopeptidase Gp63, which is involved in both parasite metabolism and infectivity (Bouvier et al. 1987). Furthermore, Leishmania glycolipids (Avila et al. 1991) could induce both anti-carbohydrate IgE and IgG antibodies, as demonstrated in schistosomiasis (van der Kleij et al. 1999). At present, we do not know the biological role played by these anti-carbohydrate IgG and IgE antibodies in the host-Leishmania relationship nor the factors influencing their production. However, the involvement of them in the interaction of Leishmania with cell receptors and possible participation in mechanisms of down regulation of leishmanial glycoprotein enzymes such as Gp63 should be investigated.
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc04112f3.jpg] [oc04112f4.jpg] [oc04112f2.jpg] [oc04112f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}