 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
| ||
|
Mem Inst Oswaldo
Cruz, Rio de Janeiro, Vol. 99, No. 6, October, 2004, pp. 545-552
Diarrheagenic Escherichia coli Categories among the Traditional Enteropathogenic E. coli O Serogroups - A Review Leila C Campos/+, Marcia R Franzolin, Luiz R Trabulsi* Departamento
de Bacteriologia, Instituto Oswaldo Cruz- Fiocruz, Av. Brasil
4365,
21045-900 Rio de Janeiro, RJ, Brasil *Laboratório Especial
de Microbiologia, Instituto Butantan, São Paulo, SP, Brasil Received
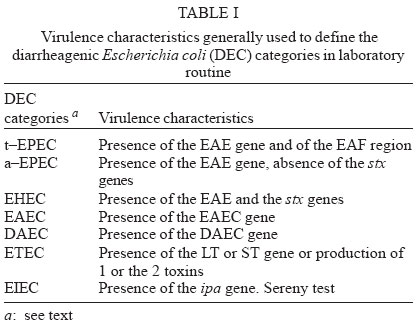
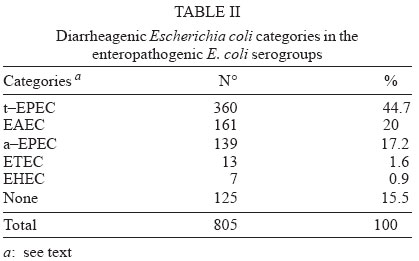
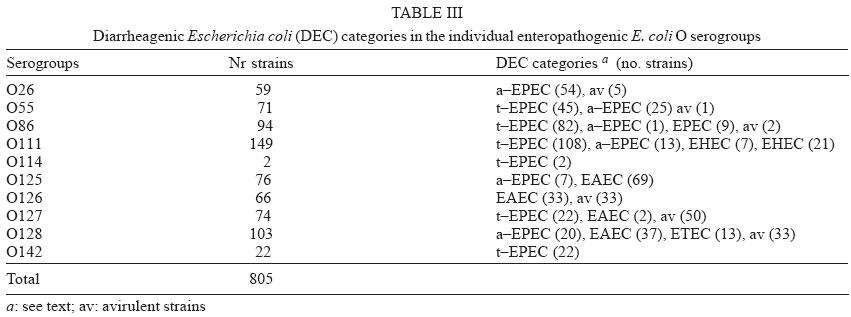
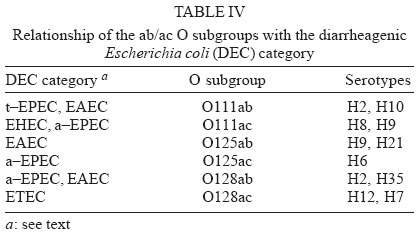
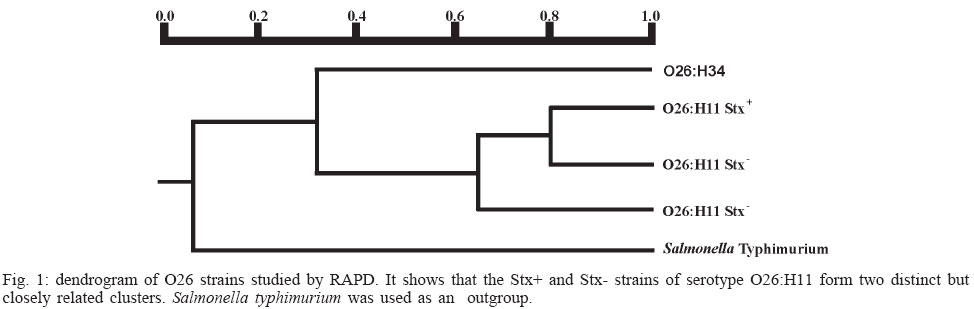
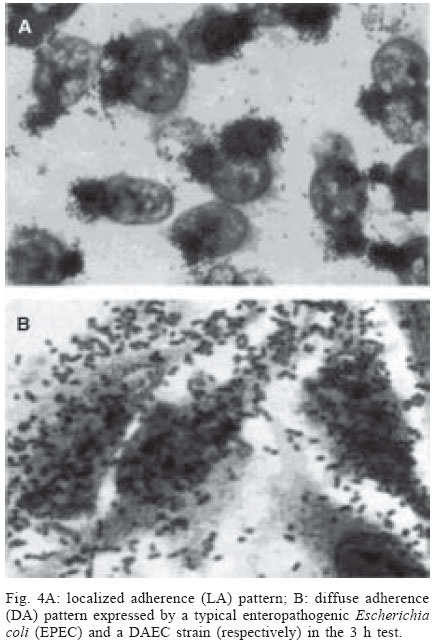
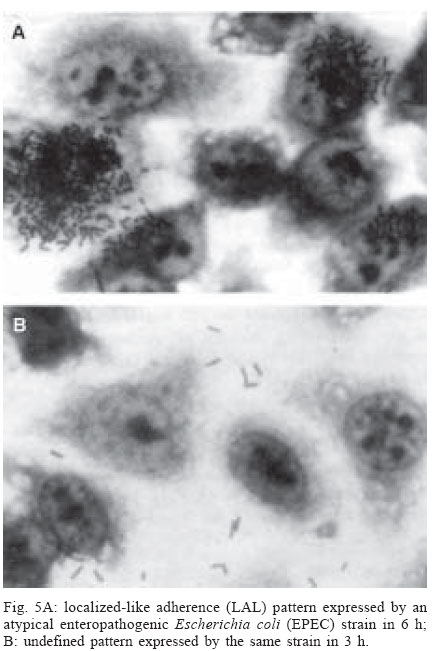
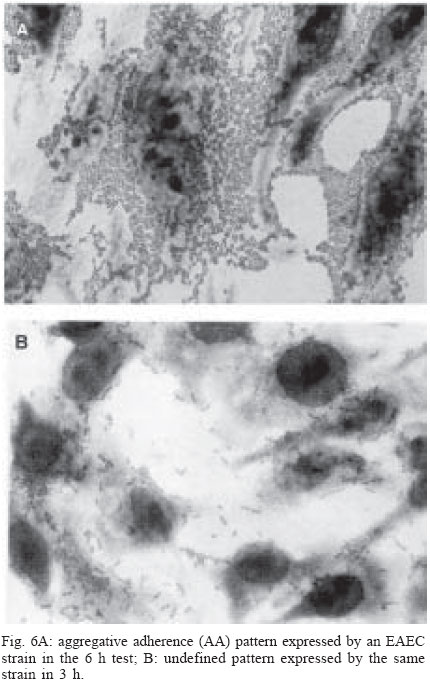
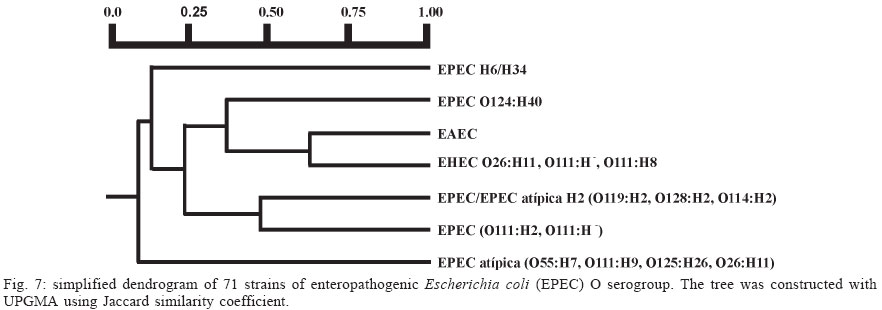
4 June 2004 Code Number: oc04116 The socalled enteropathogenic Escherichia coli (EPEC) O serogroups include typical and atypical EPEC, en-terohaemorrragic E. coli, enterotoxigenic E. coli, and enteroaggregative E. coli. The aim of this article is to review the composition of each O serogroup and the major serotypes, clones, and additional virulence characteristics of each of these diarrheagenic categories. Their adherence patterns and genetic relationships are also presented. The review is based on the study of 805 strains of serogroups O26, O55, O86, O111, O114, O119, O125, O126, O1127, O128, and O142 most of which isolated in São Paulo from children with diarrhea between 1970 and 1990. Since some O serogroups include more than one diarrheagenic category O serogrouping only should be abandoned as a diagnostic method. However serotyping is a reliable method for those serotypes that correspond to clones. Key words: diarrheagenic Escherichia coli - serogroups - serotypes - virulence factors - adhesion patterns - genetic relationship The species Escherichia coli is serologically divided in serogroups and serotypes on the basis of its antigenic composition (somatic or O antigens for serogroups and flagelar or H antigens for serotypes). Many strains express a third class of antigens (capsular or K antigens) that although important in pathogenesis only occasionally are used in serotyping. The species comprise intestinal and extraintestinal pathogens. The intestinal pathogens are also known as diarrheagenic E. coli (DEC) of which six categories have been characterized: enteropathogenic E. coli (EPEC), enterohaemorrhagic E. coli (EHEC), enterotoxigenic E. coli (ETEC), enteroinvasive E. coli (EIEC), enteroaggregative E. coli (EAEC), and diffusely adhering E. coli (DAEC) (Nataro & Kaper 1998). Recently EPEC has been divided in typical EPEC (t-EPEC) and atypical EPEC (a-EPEC) (Trabulsi et al. 2002). There are many differences between these categories but in routine they are defined by a few virulence markers (Table I). The extraintestinal pathogens (EXPEC) more frequent includes the strains associated with urinary tract infections (UPEC), neonatal meningitis (MAEC), and bacteremia. For an excellent review on DEC see Nataro and Kaper (1998) and on E. coli in general Gyles (1994). E. coli is also a very important veterinary pathogen (Gyles 1994). The term EPEC was coined by Neter et al. (1955) a half century ago to indicate the E. coli O serogroups associated with diarrhea and during many years these organisms were identified in routine by their O serogroups. As after sometime there was no consensus any more about these EPEC O serogroups the World Health Organization convened a meeting in 1987 to settle the subject. According to this meeting the following E. coli O serogroups should be considered EPEC O serogroups: O26, O55, O86, O111, O114, O119, O125, O126, O127, O128, O142, and O158 (WHO 1987). At this time already there were studies showing that at least some of these serogroups contained different diarrheagenic serotypes but their pathogenic diversity only latter became really clear (Nataro & Kaper 1998). During the last 10 years we have studied most of the EPEC O serogroups in regard to DEC categories, serotypes, clones, and genetic relationships. The purpose of this article is to review these characteristics. It is based on the study of 805 strains of serogroups O26, O55, O86, O111, O114, O119, O125, O126, O127, O128, and O142 most of which were isolated in São Paulo from children with diarrhea between 1970 and 1990. All strains were studied in regard to adhesion patterns, virulence genes implicated in the definition of the DEC category, and serotypes. The strains of serogropus O55, O111, and O119 were also studied by MLEE and a representative number of strains of serogroups O86, O127, O128, and O142 were studied by ribotyping. Serogroup O26 was studied by RAPD. Potential virulence factors were studied in a representative number of strains of the DEC categories detected (Campos et al. 1994, Rodrigues et al. 1996, Valle et al. 1997, Gonçalves et al. 1997, Monteiro-Neto et al. 1997, Dias 1998, Valle 1998, 2001, Ghilardi et al. 2003). Other studies performed will be mentioned during the review. DEC CATEGORIES IN THE EPEC O SEROGROUPS The DEC categories and the number of strains of each category as well the number of avirulent strains are shown in Table II. Typical EPEC was the most frequent category followed by EAEC, atypical EPEC, ETEC, and EHEC. EIEC and DAEC were not found among the strains tested. A number of strains showed the diffuse adherence (DA) pattern but they proved to belong to the ETEC, EAEC, and atypical EPEC categories. The number of avirulent strains was relatively high but this was serogroup dependent (see next item). The absence of the EIEC category among the EPEC O serogroups was expected because these organisms usually belong to a few other O serogroups which are not related to the EPEC O serogroups (Nataro & Kaper 1998). DAEC strains could be expected but this marker was found only in association with other well established categories. More studies are necessary to characterize DAEC as a DEC category. At present it is not possible to exclude that DA is only an unspecific marker associated with some E. coli pathotypes, including UPEC. DEC CATEGORIES IN EACH EPEC O SEROGROUP The DEC categories in each EPEC O serogroup are shown in Table III. It can be seen that the number of categories per serogroup was variable and that only serogroups O26, O126, and O142 included only one category. The remaining O serogroups included two or more categories. Serogroup O111 was the only serogroup to include four categories (typical and atypical EPEC, EAEC, and EHEC). It is interesting that EHEC occurred only in serogroup O111 and ETEC only in serogroup O128. The number of strains not belonging to any of the categories detected (avirulent strains in Table III) was relatively high in serogroups O126, O127, and O128. While the frequency of these strains was 24% in the three serogroups it was less than 2% in the remaining serogroups. We have some reasons to believe that most of these avirulent strains were virulent on isolation and that they became avirulent during storage. First we have observed that the older the strains the higher the number of avirulent ones and second during our studies we have also observed that several virulent strains were no more virulent when the tests were repeated. This was particularly frequent among the strains primarily classified as EAEC and ETEC. Interestingly enough in two EPEC serotypes of serogroups O127 and O128 there occurred several eae negative strains, a marker that only exceptionally was absent from EPEC strains from other serogroups (Dias 1998, Ghilardi et al. 2003). The occurrence of a larger number of avirulet strains in serogroups O126, O127, and O128 was also observed by Scotland et al. (1996) who examined a similar collection of O serogroups strains isolated in United Kingdom. Taken together these data suggest that these serogroups are unable to keep their virulence markers as stably as other O serogroups for reasons that are still unknown. DEC CATEGORIES AND O SUBGROUPS The O antigens of E. coli may be composed of antigenic factors that permit to divide the O serogroups in O subgroups. The common factors of these subgroups are designated "a" and the non-common "b" and "c". For example serogroup O111 is divided in the subgroups O111ab and O111ac. In our studies we managed to identify the ab and ac subgroups in serogroups O111, O125, and O128. The relationship between these O subgroups, serotypes and DEC category are shown in Table IV. It is worth to note that the ac subgroups in the 3 serogroups were the subgroups of EHEC, atypical EPEC and ETEC respectively. Most of the studies on the EPEC O subgroups were done by Ewing et al. (1963) who showed the epidemiological importance of this division. For example they showed that all O111 strains isolated from humans were of the ab subgroup while those strains isolated from monkeys were of the ac subgroup. Now we are showing that there is also a correlation between subgroups and DEC category and this could be used for diagnostic an epidemiological purposes. MAJOR DEC SEROTYPES Each DEC category included a variable number of serotypes but only 25 were considered major serotypes on the basis of their importance or frequency (Table V). It should be noted that each category has a set of specific serotypes and that among the typical EPEC serotypes only three H antigens were found: H2, H6, and H34. However the H2 and H6 antigens were common to typical and atypical EPEC. When the H antigens were identified by the restriction pattern of the fliC gene the H2 antigens of all serotypes showed a pattern identical to the H35 antigen of O subgroup 128ab (Botelho et al. 2003). This finding precludes the use of this method to identify the O128ab:H2 and O128:H35 serotypes. Under the pratical point of view this does not represent a serious identification obstacle because the two serotypes differ in several other characteristics including the adherence pattern and utilization of the phenil-propionic acid (Monteiro-Neto & Trabulsi 1999). The number of serotypes by serogroup was larger in serogroups O126, O127, and O128. Some of these serotypes could be originally avirulent and so they have contributed to the larger number of avirulent strains in these serogroups. CLONALITY OF THE MAJOR DEC SEROTYPES In general two groups of serotypes were detected: one was represented by serotypes that corresponded to single clones or to a cluster of closely related clones and the other by serotypes composed of two clones. Belonged to the first group most of the typical and atypical EPEC serotypes, the EHEC serotype, and some of the EAEC and ETEC serotypes. Belonged to the second group serotypes O26:H11, O127:H6, and O128:H2. Serotype O26:H11 showed several clones as already observed by others but the most significant aspect was the finding that Stx positive and Stx negative strains belonged to different clones (Fig. 1) (Peixoto et al. 2001). Serotype O127:H6 was composed of two quite distinct ribotypes, one with the typical EPEC markers (eae/EAF) and the other with the EAEC marker (Fig. 2) (Ghilardi et al. 2003). Serotype O128:H2 also presented two ribotypes one corresponding to atypical EPEC and the other to typical EPEC (Fig. 2) (Dias 1998). The atypical EPEC strains were isolated in Brazil and the typical one in England (Cigleris strain). We have also observed that some clones could be composed of different sero and pathotypes but the most impressive one was ET5 (serogroup O55) that included typical EPEC, atypical EPEC, and a group of strains with a combination of markers (Fig. 3) (Rodrigues et al. 1996). The study of serotypes by other methods may be helpful to understand the evolution and the epidemiology of these organisms. POTENTIAL VIRULENCE FACTORS In addition to the virulence factors that define the DEC category many others may be found among the O serogroup strains. So far a role for these factors in pathogenesis has not been established. In our studies the most frequent of these factors were Hly (enterohemolysin) in the EHEC strains (Campos et al. 1994), EAST1 (enteroaggregative heat-stable enterotoxin 1) in EHEC and atypical EPEC (Trabulsi et al. 2002) and CDT (cytolethal distending toxin) in all O86:H34 strains (Ghilardi et al. 2001). In adition the EAEC strains showed the factors included in Table VI (Elias et al. 2002). ADHERENCE PATTERNS Most typical EPEC strains showed the localized adherence (LA) pattern (Fig. 4a) and this pattern was associated with the expression of bundle-forming pilus (BFP) in all strains tested (Gismero-Ordonez et al. 2002). This pattern could be used to define t-EPEC since it was regularly expressed and specific. Only exceptionally it was not expressed by the eae/EAF positive strains in the 3 h test. The localized-like adherence (LAL) pattern (Fig. 5a) was characteristic of the EHEC and atypical EPEC strains but it was fully expressed only after 6 h of incubation. In the 3 h test most strains exhibited an undefined pattern (Fig. 5b). The LAL pattern was partially mediated by intimin (Pelayo et al. 1999). The EAEC category was defined by the aggregative adherence (AA) pattern (Fig. 6a) since a relatively large proportion of strains did not react with the EAEC probe. Although defining EAEC, the AA pattern was expressed by all O125ac:H6 strains which we have classified as atypical EPEC (Valle et al. 1997, Trabulsi et al. 2002). In many strains the AA pattern is expressed only after 6 hours of incubation (Fig. 6a). As said before strains of the DAEC category was not identified in this study but the DA pattern was expressed by a few atypical EPEC, ETEC and EAEC strains. In the atypical EPEC strains the DA pattern (Fig. 4b) was mediated by the AfaE protein (Keller et al. 2002). GENETIC RELATIONSHIPS The dendrogram in Fig. 7 illustrates the genetic similarity and degree of divergence between t-EPEC, a-EPEC, EHEC, EAEC, and their serotypes. The more accentuated divergence occurred between the typical EPEC with H6/H34 antigens (cluster A) and the a-EPEC of serotypes O26:H11, O55:H7, O111:H9, and O125ac:H6 (cluster C). The other categories and respective serotypes occupied an intermediary position (cluster B) where two sub-clusters (B1 and B2) are proeminent. B1 includes a t-EPEC serotype (O127:H40) and all EAEC, and EHEC strains and B2 all typical and atypical EPEC strains with the H2 antigen. In subcluster B1 typical EPEC, EAEC and EHEC can be clearly differentiated and the same is true for the O111:H2 and the other H2 serotypes in cluster B2. The dendrogram suggests that the similarity and the divergence seen is dictated both by the DEC category and by the serotype. The genetic relationships of the EPEC O serogroups have been studied by several authors, with special emphasis by Wittham & McGraw (1996). These authors have shown that the EPEC serotypes can be divided in two clonal groups named EPEC1 and EPEC2 and that the first group express the H6 and the H34 antigens and the second the H2 antigens. Essentially these results are identical to the ones reported here. They also have shown that the EHEC strains of serogroups O26 and O111 clusters together and are genetically different from serotype O55:H7. In their studies EAEC strains were not included but it is demonstrated here that they are different but closely related to the EHEC strains It is not known why some O serogroups are more efficient to receive or maintain determined virulence factors and so far as we know no studies have been made to clarify this. COMMENTS The occurrence of the DEC categories in the EPEC O serogroups has been investigated by several authors (Blanco et al. 1993, Giammanco et al. 1996, Paciorek 2002). One of the most complete study was performed by Scotland et al. (1996) in United Kingdom. These authors examined around 800 E. coli strains belonging to the same O serogroups examined by us in regard to serotypes and the genes that define the DEC categories with exception of the genes for ETEC. The results obtained are similar to ours in the sense that they have detected the same DEC categories (with exception of ETEC, of course) and most of the serotypes identified in our studies. The most significant differences were in respect to the frequency of the categories and the kind of serotypes. For example, the most frequent categories in their studies were a-EPEC, EAEC, and EHEC while in our studies the most frequent ones were t-EPEC and EAEC. Regarding serotypes, it was surprising the total absence in UK of the most frequent t-EPEC serotypes in our studies, namely O111:H2 and O55:H6. In the past these two serotypes were very frequent in UK as a cause of diarrhea outbreaks and sporadic cases of infantile diarrhea. Comparison of the frequency of EPEC and EHEC along the years in UK (and others developed countries) and in Brazil offers some interesting aspects. In the past the EPEC was extremely frequent in UK (Smith et al. 1996) and others developed countries and EHEC was practically inexistent. At present t-EPEC are very rare and the atypical EPEC and EHEC are frequents. In Brazil (São Paulo) we need to consider two periods. Until the end of the 1980s, t-EPEC was the predominant agent of diarrhea and EHEC extremely rare (Toledo et al. 1983, Gomes et al. 1989, 1991). In fact at that time we had isolated only three O111ac:H- strains (Stx+) and a very few strains of others serotypes (Campos et al. 1994). At present this situation has changed substantially. It seens that we are approaching the present situation in developed countries which was started in the 1960s for t-EPEC and the 1980s for EHEC. Accordingly most of our EPEC isolates lack EAF and we have started to isolate O157:H7 and O26:H11 Stx+ strains. The reasons for this kind of events have not been established but they are not exclusive of this pathogen. They are similar to the events that have occurred with Salmonella serotypes: a dramatic fall in the frequency of S. Typhi and a vast increase in the frequency of S. Typhimurium and similar serotypes. S. Typhi as typical EPEC is specific human pathogens and S. Typhimurium as EHEC and atypical EPEC are pathogens associated with humans and animals. The reductions of the typical EPEC frequency in Brazil (São Paulo) may be related to the improvement of the living conditions in recent years. Others studies on DEC categories in the EPEC O serogroups were performed by Beutin et al. (1990) in Germany and Yam et al. (1994) in Australia. Beutin et al. (1990) studied a large number of O114 strains in regard to serotypes, electrophoretic types, and virulence factors. They have detected t-EPEC, ETEC and probably UPEC. There was strong correlation between serotypes, clones and pathogens categories. Yam et al. (1994) studied a group of O126 strains and detected ETEC, and EAEC and found a good correlation between serotypes, clones and virulence characteristics. There are many others studies on Stx producing strains of serotypes O26:H11 and O111ac:H-. From the practical point of view it is evident that serogrouping (identification of the O antigen only) is not a reliable approach to identify EPEC as it still done in certain laboratories. However serotyping is entirely reliable but only for those serotypes that correspond to clones. ACKNOWLEDGMENTS To Silvia Bando for constructing the dendrograms in Figs 1 and 7. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc04116f6.jpg] [oc04116t1.jpg] [oc04116t3.jpg] [oc04116f3.jpg] [oc04116f2.jpg] [oc04116t4.jpg] [oc04116t5.jpg] [oc04116f1.jpg] [oc04116f4.jpg] [oc04116t6.jpg] [oc04116t2.jpg] [oc04116f5.jpg] [oc04116f7.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}