 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 6, October, 2004, pp. 597-601 Biology of the First Generation of a Laboratory Colony of Nyssomyia intermedia (Lutz & Neiva, 1912) and Nyssomyia neivai (Pinto, 1926) (Diptera: Psychodidae) José Dilermando Andrade Filho/+, Eunice A Bianchi Galati*, Alda Lima Falcão Laboratório
de Leishmanioses, Centro de Pesquisas René Rachou-Fiocruz, Av. Augusto
de Lima 1715, 30190-002 Belo Horizonte, MG, Brasil *Departamento de Epidemiologia,
Faculdade de Saúde Pública, Universidade de São Paulo,
São Paulo, SP, Brasil Financial support: CNPq, Fiocruz Received 1 June
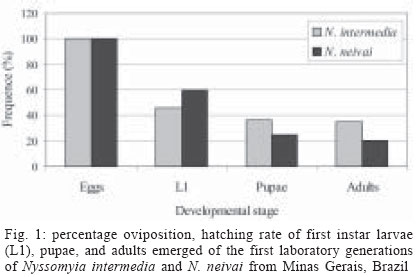
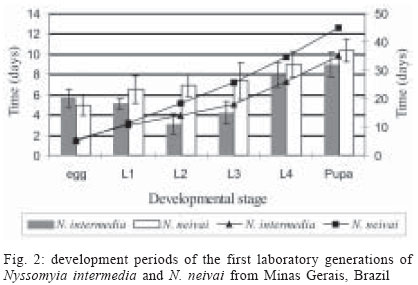
2004 Code Number: oc04126 The phlebotomine sand flies Nyssomyia intermedia (Lutz & Neiva, 1912) and Nyssomyia neivai (Pinto, 1926) are very close and may be involved in the transmission of Leishmania spp. Ross, 1903 in Brazil. The biology of the first laboratory-reared generations of these species, descended from insects captured in Além Paraíba (N. intermedia) and Corinto (N. neivai) in the Brazilian state of Minas Gerais, is described here. The captured females were fed on hamsters and maintained individually in rearing pots. Laboratory temperature and relative humidity were maintained at 25-26oC and 80% respectively. The productivity of the first generation of N. intermedia was greater than that of N. neivai, and its development time clearly shorter, particularly for the second and third larval instars. Key words: Nyssomyia intermedia - Nyssomyia neivai - Phlebotominae - Psychodidae The phlebotomine sand flies Nyssomyia intermedia (Lutz & Neiva, 1912) and Nyssomyia neivai (Pinto, 1926) are very closely related species considered to be conspecific until about 10 years ago (Martins et al. 1978, Young & Duncan 1994). Males from both species can be distinguished by means of the genital filament (Andrade Filho et al. 2003) while females may be recognized by spermathecal characters such as length, number of annulae, shape and width of spermatheca head, and length of common duct in relation to individual duct (Marcondes 1996, Andrade Filho et al. 2003). Both N. intermedia and N. neivai may be involved in the transmission of the Leishmania spp. Ross, 1903 causing American cutaneous leishmaniasis (ACL), particularly in Southeastern (SE) Brazil. Silva and Gomes (2001) and Gontijo et al. (2002) dissected several females of N. intermedia sensu lato and N. intermedia sensu stricto, respectively from different ACL foci but did not detect Leishmania. Both species bite humans avidly (Oliveira Neto et al. 1988, Gomes et al. 1989) and although N. intermedia s.l. has been found naturally infected with flagellates in SE Brazil (Forattini et al. 1972, Rangel et al. 1984, Casanova et al. 1995) and predominates among man-biting sand flies in several endemic or epidemic foci of ACL (Rangel et al. 1990, Gomes et al. 1995, Salomón et al. 2001, Gontijo et al. 2002), it is still only regarded as a suspected vector (Killick-Kendrick 1990), based on the fact that conspecific parasites have not been identified from isolates obtained from humans and dogs. Laboratory colonization of phlebotomine species is very important for studies on their biology, particularly their interactions with Leishmania and several other pathogens (Coelho 1962, Coelho et al. 1967, Killick-Kendrick et al. 1977, Rangel et al. 1992). Species that have been successfully reared in the laboratory include N. intermedia s.l. (Rangel et al. 1985, 1986, Brazil et al. 1997, Ferro et al. 1998, Liutgards-Moura et al. 2000, Alves et al. 2000). In this paper we compare the biology in the laboratory of the first generation of N. intermedia and N. neivai. MATERIAL AND METHOD The method for rearing of the two species were based on Killick-Kendrick et al. (1977) and Modi and Tesh (1983). The classification scheme used is that of Galati (2003). Specimens of N. intermedia were collected on 27- 28 August 2002 in an area known as Benjamin Constant, Além Paraíba, state of Minas Gerais (21o53'S; 42o42'W) from where Lutz and Neiva (1912) described this species. This area still retains some of the original Atlantic Forest, although cattle are grazed in clearings. Several investigators have collected sand flies in this area (Rangel et al. 1985, Marcondes 1997, Falcão pers. inform.) where N. intermedia is abundant. The light traps (Falcão 1981) used for sampling were hung in a cattle pen, a pigsty, a chicken house in a mango tree (used as a roosting-place by the birds) and a kennel used by a dog at night. These traps were left in place from 18:00-07:00 h. A white illuminated Shannon trap (Shannon 1939) was also hung in a peri-domestic habitat of the Barra do Ouro Fino farm where sand flies were collected, by direct aspiration, from 18:00-22:00 h. Specimens of N. neivai were collected in Corinto, also in the state of Minas Gerais. This collection site (17o58'S; 49o32'W) is approximately 8 km from the town of Lassance. There are several buildings in the area that are used by local residents and fishermen, fishing and agriculture being as the main economic activities of the region. The predominant vegetation type is cerrado. Sand flies were sampled using an illuminated Shannon trap in a chicken house localized in Lavado region, about 3 m from Rio das Velhas on July 4, 2002 from 18:00-23:00 h, this being the period of peak activity of N. neivai (data provided by Centro de Referência Nacional e Internacional para Flebotomíneos do Centro de Pesquisas René Rachou-Fiocruz) After recovering the traps, sand flies were transferred to 11 cm diameter rearing containers, each lined with plaster to maintain humidity and with a hole in the base plugged with plaster, that could be placed on a damp substrate. The containers were topped with nylon mesh with a hole cut in the centre, through which sand flies were introduced using a Castro aspirator (Castro pers. inform. by Barretto & Coutinho 1940 apud Pessoa & Barretto 1948). When not in use the hole was plugged with cotton wool. A cotton wool swab soaked in sugar solution was placed on the mesh as an energy source. Sand flies were fed in the field on hamsters [Mesocricetus auratus (Water House, 1839)] anaesthetised with 0.7-0.8 ml Thiopental (1 g/20 ml distilled water) and placed directly on top of the containers, allowing sand flies to feed through the mesh. Rearing containers containing the sand flies were transported from the collection site to the laboratory in Tupperware© boxes lined with moistened paper towels. In the laboratory, the sand flies were allowed to feed on hamsters a second time. Females that survived 48 h after blood feeding were then transferred to individual (oviposition) vials 4 cm in diameter. Cotton wool swabs soaked in fresh sugar solution were placed on the mesh of the oviposition vials. The vials were inspected daily to check whether the females had laid eggs. Females that had oviposited and were still alive were given the opportunity to take a new blood meal from a hamster, Japanese quail (Coturnix coturnix japonica Temminck and Schlegel, 1849) or human volunteer. After each female sand fly died, it was mounted in Berlese fluid on a microscope slide labelled with the same number as the oviposition vial. These preparations were examined under the microscope for species identification. Progeny of the wild-caught insects were reared in the laboratory at 25-26oC and relative humidity of approximately 80%. The oviposition vials containing eggs were monitored daily and after larvae emerged they were fed with a powdered diet containing equal proportions of "xaxim" (an arboreal Pteridophyta), dry rabbit faeces, dehydrated lettuce, and Vitormonio© fish food. The vials were examined daily to monitor possible contamination by fungi or mites and to remove excess faeces or leftover food. The numbers of eggs per female, larvae in each stage, pupae, and adults were noted as development progressed. Developmental parameters of N. intermedia and N. neivai such as incubation periods, duration of larval stages and adult eclosion were calculated as averages of times from the first to the final day of the stage in question. RESULTS Data obtained for N. intermedia and N. neivai concerning sex ratios, productivity and development times are compared in Figs 1 and 2. Nyssomyia intermedia - Field-collected females fed actively on hamsters and also tried to bite man although they were not allowed to do so. A total of 123 females collected in Além Paraíba were separated into individual oviposition/rearing chambers and 72 of these (58.5%) laid eggs. A single female that did not belong to either of the two species being studied was identified as Nyssomyia whitmani (Antunes & Coutinho, 1939). The percentage of N. intermedia females that laid eggs was 57.7% and eggs hatched in all of the rearing chambers. Thirty-nine (54.2%) of the females that laid eggs survived oviposition but none of them took a second blood meal. The total number of eggs laid was 3228, a mean of 45.5 per female (range 12-78). From these 1474 first instar (L1) larvae were obtained, representing a hatch rate of 45.7%. A total of 1144 adults emerged (35.4% of the eggs and 77.6% of larvae hatched) in 60/72 (82.3%) of the rearing chambers where oviposition occurred of these 51.5% were females and 48.5% males (M:F sex ratio 0.94:1). Male sand flies were the first to emerge. The entire egg to adult period ranged from 29 to 42 days (average 34.9 days) for N. intermedia. Average duration of the egg stage was 5.7 days, while duration of each larval stage (in days) was as follows: L1 5.1; L2 3.1; L3 4.2, and L4 7.9. The pupal stage was the most prolonged, lasting a mean of 9.0 days. Nyssomyia neivai - As with N. intermedia, N. neivai females fed readily on hamsters. They also bit man, but not japonica quail, when these were offered as feeding sources. A total of 91 females collected in Corinto were separated into individual oviposition/rearing chambers. One of the 54 N. neivai rearing chambers was found to contain a different species, identified as Evandromyia lenti (Mangabeira, 1938). Only eight (15.1%) of the remaining 53 females survived oviposition and these did not take another blood meal. A total of 2573 eggs was laid by the 53 N. neivai females, a mean of 48.5 per female, with up to 70 eggs in a single batch. The 2573 eggs produced 1552 L1 larvae, a hatch rate of 60.3%. Adult sand flies were produced from 39/53 (73.6%) of the oviposition/rearing chambers. Adult survival rate as a proportion of eggs laid was 533/2573 (20.7%) while 533/1552 (34.3%) of L1 larvae completed development. The M:F sex ratio was 43.3/56.7% (0.76:1) Males were the first to emerge, as in N. intermedia. The entire egg to adult period for N. neivai was 45.1 days (range 36-54). Average duration for the egg stage was 5 days. There was little variation between any of the first three larval stages: the L1 lasted a mean of 6.5 days, L2 6.9, and L3 7.3. The L4 and pupal stages lasted a mean of 9.0, and 10.4 days respectively. DISCUSSION The fact that 99% of all sand flies collected in Benjamin Constant and Corinto were either N. intermedia or N. neivai indicates that these areas may be used to provide more material for laboratory rearing studies in the future. Under laboratory conditions, N. intermedia females remained alive after oviposition, unlike those of N. neivai, which had a low survival rate. This suggests that the former species would be more important in Leishmania transmission, since it would be able to take a second blood meal. The fact that none of the insects re-fed in the laboratory, even when offered the chance to bite hamsters, japonica quail or humans, may have been due to the fact that they were held individually, and thus were isolated from males or other females whose presence might stimulate blood feeding. Gomes et al. (1982) reported that N. intermedia s.l. fed twice under natural conditions. This was also observed by Brazil et al. (1991) for N. intermedia and Dia-Eldin et al. (1992) for Lutzomyia longipalpis (Lutz and Neiva, 1912), both in nature and in the laboratory. In the wild, N. intermedia has also been reported to survive after oviposition (Gomes et al. 1982). Sand fly survival is rare under laboratory conditions, the few records involving very low rates, including approximately 17.7% for Lutzomyia renei (Martins Falcão & Silva, 1957) (Alves et al. 2001) and 8.3% for L. longipalpis (Souza et al. 1995). According to Casa-nova (2001), N. neivai would be able to feed up to three times within 10 days. The percentage of individualized females that oviposited was nearly the same for both species under study, at 57.5% for N. intermedia and 59.3% for N. neivai. Such values correspond closely with those reported by Rangel et al. (1985), who worked with four generations of N. intermedia, in which 58.9% of blood-fed females laid eggs. Casanova (2001) found that N. neivai fed mostly on human blood in peridomiciliary areas, although he considered this species to be opportunistic rather than truly anthropophilic, feeding preference varying with host availability. Aguiar et al. (1987) arrived at similar conclusions for N. intermedia in Itaguaí in the Brazilian state of Rio de Janeiro. According to Rangel et al. (1986), the number of N. intermedia females that laid eggs and the number of eggs per female depend on the blood source. The mean number of eggs laid per female was quite similar for both species, although slightly higher for N. neivai. The means found here are higher than those observed by Rangel et al. (1985), who recorded 40.2 eggs per N. intermedia female in the first generation, or by Alves et al. (2001) who reported 15.7 eggs per female in field-collected L. renei. The mean was also significantly higher than the 32.7 eggs per female observed for Evandromyia carmelinoi (Ryan, Fraiha, Lainson, Shaw, 1986) (Alves et al. 2000). During the present study the hatch rate in N. neivai was 60.3%, much higher than that of N. intermedia (45.7%). Similar values were found for N. intermedia by Rangel et al. (1985), i.e., 64.1%. Values for both species are much higher than those of L. renei, which showed a hatching rate of 24% (Alves et al. 2001). The productivity of N. intermedia was higher than that of N. neivai, the percentage of eggs that ultimately gave rise to adults in the former being 35.4%. In all, 77.6% of the L1 larvae reached the adult stage, representing a very low mortality rate during development. The corresponding values for N. neivai were much lower, at 20.7% and 34.3% respectively. It is noteworthy that the highest mortality rate among phlebotomine sand flies occurs between the first and second developing stages and that these developmental stages were more prolonged in N. neivai than in N. intermedia, particularly L2. This might have affected the mortality rate at this stage. In L. renei only 8.6% of the eggs and 35.6% of the L1 larvae ultimately gave rise to adults (Alves et al. 2001). Rangel et al. (1986) recorded much higher indices for N. intermedia at 34.4% and 53.6% respectively. The M:F ratio was higher for N. intermedia, although Rangel et al. (1985) found that 45.1% of the emerging adults of this species in the laboratory were females. Males emerged before females, as has been reported by other authors (Chaniotis & Anderson 1964, Killick-Kendrick et al. 1977, Montoya-Lerma et al. 1998, Alves et al. 2000). N. neivai had a longer life cycle than N. intermedia, the two species requiring 45 and 35 days respectively to complete development. This fact might be due to the longer developing period of second and third instar larvae, an average of 7 days for each stage, whereas for N. intermedia the media was 7 days to complete both phases (3 days for L2 and 4 for L3). Developing periods for immature stages of both species have been shown to be higher than for L. longipalpis, lasting 29.5 days (Brazil et al. 1997). According to Rangel et al. (1985) the entire cycle for the first generation of N. intermedia lasted 37 days, very close to the results of the present study and a little longer than the 32.8 days reported by Brazil et al. (1997). The development period of N. neivai was close to that of E. lenti, which requires 40.2 days (Brazil et al. 1997). The longest developmental stages were the L4 and pupa, both of which involve more radical physiological changes and tissue formation than previous instars (Forattini 1973). Several physical factors may affect phlebotomine biological cycles (Rangel et al. 1987, Nieves et al. 1997), besides biological factors such as larval diet (Rangel et al. 1985, Wermelinger & Zanúncio 2001). Attempts during the present study to maintain colonies of N. intermedia and N. neivai in larger rearing chambers were unsuccessful, since F2 females died just after taking blood meals. It may be that symbiotic protozoa or bacteria, possibly acquired from natural diet during larval instars or sugar meals of females, assist in bloodmeal digestion. These are not present in the larval diet or sugar meals provided to adult females in the laboratory and this may explain the low survival rate of the insects. Studies should be performed on intestinal microbiota of wild-caught N. intermedia and N. neivai in order to provide further information on this aspect of their biology, as has already been done for L. longipalpis (Oliveira et al. 1999, 2001).
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc04126f2.jpg] [oc04126f1.jpg] |
| |||||||||

{kind=link}
{kind=link}