 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
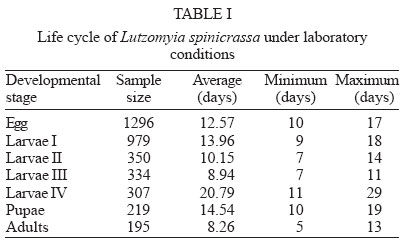
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 6, October, 2004, pp. 603-607 Life Tables and Reproductive Parameters of Lutzomyia spinicrassa (Diptera: Psychodidae) under Laboratory Conditions Jesús Escovar, Felio J Bello/+, Alberto Morales, Ligia Moncada*, Estrella Cárdenas Laboratorio de
Entomología, Biología Celular y Genética, Departamento
de Ciencias Básicas, Universidad de La Salle, Bogotá DC, Colombia *Laboratorio
de Parasitología, Facultad de Medicina, Universidad Nacional de Colombia,
Bogotá DC, Colombia Financial support: Colciencias (grant 1243-05-12416), Universidad de La Salle, Bogotá Colombia Received 14 June
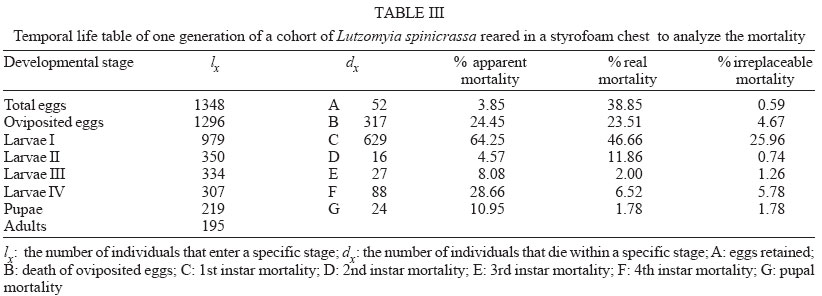
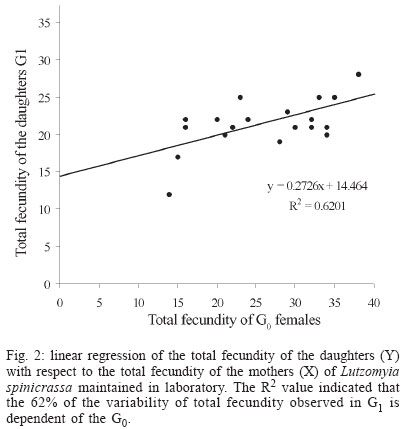
2004 Code Number: oc04127 Lutzomyia spinicrassa is a vector of Leishmania braziliensis in Colombia. This sand fly has a broad geographical distribution in Colombia and Venezuela and it is found mainly in coffee plantations. Baseline biological growth data of L. spinicrassa were obtained under experimental laboratory conditions. The development time from egg to adult ranged from 59 to 121 days, with 12.74 weeks in average. Based on cohorts of 100 females, horizontal life table was constructed. The following predictive parameters were obtained: net rate of reproduction (8.4 females per cohort female), generation time (12.74 weeks), intrinsic rate of population increase (0.17), and finite rate of population increment (1.18). The reproductive value for each class age of the cohort females was calculated. Vertical life tables were elaborated and mortality was described for the generation obtained of the field cohort. In addition, for two successive generations, additive variance and heritability for fecundity were estimated. Key words: Lutzomyia spinicrassa - life cycle - reproduction - population - heritability The sand fly Lutzomyia spinicrassa (Morales, Osorno-Mesa, Osorno & de Hoyos, 1969) belongs to the group verrucarum, series townsendi and it has a wide geographical distribution in Colombia and Venezuela, mainly in coffee plantations. In Colombia, this species is vector of Leishmania braziliensis (Young et al. 1987, WHO 1990) and has been found around plantations of coffee in several towns of Boyacá, Norte de Santander, and Sucre departments (Alexander et al. 2001, Bejarano et al. 2003). Maingon et al. (1994) founded high populations of L. spinicrassa in coffee plantations infected with two species of Leishmania: L. mexicana and L. braziliensis in the states of Trujillo, Mérida, and Táchira (Venezuela). It is an antropophilic species and their feeding activity carries out mainly in the twilight and night hours, during every month of the year. The adults rest during the day inside tree holes (Alexander et al. 1992). The sand fly demands care and special maintenance for the establishment of a colony in the laboratory. Killick-Kendrick et al. (1977) were successful for the first time on colonizing and reproducing L. longipalpis (Lutz & Neiva). Their work was the basis for the establishment in Colombia of colonies of other species of Lutzomyia, such as those achieved by the following authors: Morales et al. (1984) with L. walkeri, Neira et al. (1998) with L. torvida and L. longiflocosa, Montoya et al. (1998) with L. evansi, Ferro et al. (1998) and Cárdenas et al. (1999) with L. shannoni, Cabrera et al. (1999) with L. ovallesi and Cabrera and Ferro (2000) with three species of Lutzomyia of the group verrucarum. The reproductive cycles of arthropod vectors play a fundamental role in the epidemiology of the disease they transmit. Parameters affecting transmission of disease include fecundity rate, mortality rate, density, distribution by ages, migration rate, and genetic variation of the vector arthropods (Black & Moore 1996, Tabachnick & Black 1996). However, none of this information is available for L. spinicrassa. To better understand baseline parameters of the life cycle, growth under experimental conditions was monitored. Horizontal life table was constructed based on the developmental time of each instar in a cohort of 100 females. Additionally, predictive population para-meters were calculated. Stage specific mortality data were collected and for two successive generations, the additive variance and heritability of fecundity were estimated. MATERIALS AND METHODS Sampling information - Adults of sand flies were collected during the night (19-22 h) by aspiration using a Shannon trap in a traditional coffee plantation at 1700 m elevation near Guateque, Department of Boyacá (4º1'N, 73º30'W) on the Cordillera Oriental, Colombia. Following capture, the sand flies were transferred to cloth cages (20 cm x 20 cm) and the females were fed on hamster blood as follows: a hamster was anesthetized (sodium pentothal, 10 mg/kg animal weight) and introduced into the cloth cages for 1 h. After blood feeding, 100 couples were separated into 100 rearing chambers (5.5 cm high and 6.6 cm in diameter) (Ferro et al. 1998) and the remainder of the sand flies were introduced in rearing chambers each with 50 sand flies. After blood feeding, a 60% glucose solution was provided daily on a cotton swab placed on top of the muslin covering of each rearing chamber. The rearing chambers were introduced in styrofoam chests to be transported to the Entomology Laboratory of the Universidad de La Salle, Bogotá. Maintenance of sand flies in the laboratory - The cohort of 100 couples and the remaining adults of L. spinicrassa captured in the field were maintained in styrofoam chests in the Entomology Laboratory of the Universidad de La Salle, Bogotá. Rearing and feeding of adults and larvae of L. spinicrassa followed the methods of Modi and Tesh (1983), Endris et al. (1982), and Ferro et al. (1998). Laboratory conditions recorded during the study were as follows: average relative humidity of 85%, average temperature of 20°C. Data from field cohort (100 couples) - Each day, data were collected regarding blood feeding, date of oviposition, number of oviposited eggs, number of retained eggs, and number of days that each female survived in the adult stage. Life cycle of L. spinicrassa was estimated by daily monitoring of the development of eggs oviposited by cohort females. The total fecundity was estimated by adding the eggs oviposited to those retained by cohort females. Retained eggs were counted using post-mortem dissection of each cohort female. Data for G1 cohort (100 couples) - The cohort consisted of 100 couples (one female and one male) taken from the first generation of the field females. Immediately after blood feeding, each couple was separated and placed into a rearing chamber (5.5 cm high and 6.6 cm in diameter). The chambers were placed in plastic boxes with lids, and temperature and humidity were monitored within each box. The plastic boxes were maintained inside in styrofoam chests. Each day, data were collected on the number of live females, date of emergence of the pupae, date of blood feeding, date of oviposition, number of oviposited eggs, number of retained eggs per female, and number of days that each female survived in the adult stage. Estimation of parameters based on horizontal life tables - A horizontal life table was constructed with the data of the G1 cohort and population predicted and observed parameters were estimated using the methods developed by Southwood (1978), Rabinovich (1980), Pianka (1988), and Cárdenas et al. (1999). Vertical life table and estimation of mortality - A vertical or temporal life table was constructed based on developmental stages of G1. The apparent mortality, real mortality and irreplaceable mortality were estimated following to Southwood (1978), and Cárdenas et al. (1999). Estimation of
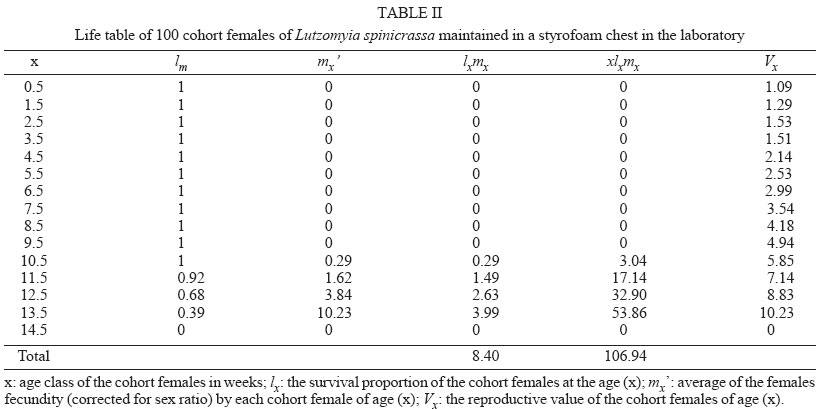
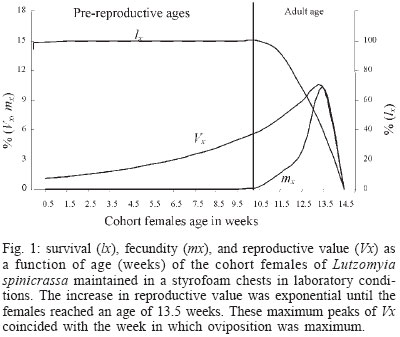
genetic parameters - Genetic parameters such as additive variance ( RESULTS Life cycle - The developmental time from egg to adult ranged from 59 to 121 days, with 12.74 weeks in average. Table I shows data for the life cycle for L. spinicrassa obtained in styrofoam chests under laboratory conditions. Horizontal life table and predictive population parameters - Table II shows the pre-reproductive age average of cohort females (G1). This value was in average 9.5 weeks and the value for reproductive age averaged 4 weeks. The pre-reproductive data provided an estimate of reproductive value (Vx). The mx , column was formed according to Rabinovich (1972). The predictive population parameters were estimated in absence of mortality according to Southwood (1978). The following values were obtained from Table II for predictive population parameters: net reproductive rate Ro = 8.4 daughter females per cohort female, generation time Tc = 12.74 weeks, the natural rate of increase rm = 0.17 daughter females for each cohort female per week, and the finite rate of increase for each cohort was l = 1.18 individuals per female per week. The reproductive values per age class of the cohort females were estimated. The reproductive value (Vx), fecundity (mx) and cohort females survival (lx) versus cohort females age (x) were plotted (Fig. 1). The increase in reproductive value was exponential (despite a fecundity of zero between 0.5 to 9.5 weeks) until the females reached an age of 13.5 weeks. These maximum peaks of Vx coincided with the week in which oviposition was maximum. Vx then abruptly decreased in the following week when the majority of the cohort females died. This population parameter provided a clear estimate of the population increase potential of L. spinicrassa in laboratory conditions. Analysis of stage-specific mortality - Vertical life table was constructed based on the lx (live individuals) and dx (mortality) at each stage of the first generation. Stage-specific mortality was based on the data obtained from the vertical life tables according to Southwood (1978) (Table III). Egg mortality - Egg mortality occurred in two levels: the first was due to retained eggs by the cohort females, whose real mortality was 3.85%. The second level consisted of oviposited by the cohort females, whose real mortality was 24.45% (Table III). Larval mortality - Significant mortality occurred during the first-instar larval (64.25%) (Table III). The conditions in the styrofoam chest environment were not favorable for the 1st-instar larval survival. The mortality at 4th-larval instar was probably due to bacterial infection. We noted that affected larvae fed at a lower rate, demonstrated a loss of mobility, and became progressively darkened. Pupal mortality - Mortality at this stage was moderate (10.95%), with the percentage developing to adults exceeding 89% (Table III). Irreplaceable mortality - The highest irreplaceable mortality (25.96%) occurred in 1st instar larvae of L. spinicrassa maintained in styrofoam chest in the laboratory (Table III). Without this mortality, an additional 629 larvae will have been available for a total of 979 2nd instar larvae. The second highest irreplaceable mortality (5.78%) occurred in the 4th-instar larvae. Without this mortality, an additional 88 larvae will have available for a total of 307 pupae. Analysis of fecundity heritability - In the previous analysis, the potential population increment was estimated, based on the production of female daughters per cohort female. To determine how much of the fecundity potential is hereditary and how much due to environment, the number of total eggs was recorded for each of the 36 females and compared with the average of the total number of eggs of their daughters. Linear regression for the total fecundity is shown in Fig. 2. An F test was applied to the regression coefficient for total fecundity; the null hypothesis was rejected at the levels 95% and 99%. Therefore, the observed variability in total fecundity was due to heritable factors. The R2 value indicated that the 62% of the variability of total fecundity observed in G1 is dependent of the G0. DISCUSSION Maintenance of laboratory colonies of phlebotomine sand flies is necessary to investigate several aspects of these vectors, such as pathogen transmission, population genetic comparisons, systematics and insecticide susceptibility among others. However, due to the great difficulty of maintaining most sand fly colonies for longer than a few generations, life-table studies on sand flies are scarcest. For New World sand flies, the studies of Killick-Kendrick et al. (1977) on L. longipalpis (Lutz & Neiva) and Cárdenas et al. (1999) on L. shannoni (Dyar) have been focused on development times, reproductive rate, life tables and reproductive parameters; whereas the studies of Morales et al. (1984), Neira et al. (1998), Montoya et al. (1998), Ferro et al. (1998), Cabrera et al. (1999), and Cabrera and Ferro (2000) have been focused on life cycle and development time of several species of Lutzomyia from Colombia. Data on life cycle and reproductive biology for phlebotomines are nearly impossible to obtain in the field because under these conditions, the immature stages have not been found. The developmental time of L. spinicrassa from egg to adult was ranged from 59 to 121 days, with 12.74 weeks in average. The sames species maintained in an incubator (temperature ranged from 25ºC to 27ºC) developed from 58 to 78 days, with 11 weeks in average (Cabrera & Ferro 2000). These findings suggest that the developmental time was influenced by the temperature. Improvement of the rearing practices is needed for an efficient establishment of a L. spinicrassa colony in the laboratory. Maintenance of suitable conditions such as temperature and humidity have been previously proven to be critical in incubation chambers for the maintenance of sand fly laboratory colonies (Killick-Kendrick et al. 1977, Morales et al. 1984, Neira et al. 1998, Montoya et al. 1998, Cárdenas et al. 1999, Cabrera & Ferro 2000). The heritability
for the total fecundity of L. spinicrassa was 56%, whereas in L.
shannoni, the heritability for total fecundity was 77.8% (Cárdenas
et al. 1999). Strickberger (1978) considered that heritability is low for
characters essential for survival, such as the rapidity of conception or
size of litter because if this value is incremented, the survival is decreased.
The additive variance for total fecundity was The high heritability of the total fecundity is reflected in the maximum intrinsic rate of population increase (rm), and the data support the assertions of Southwood (1978) that this parameter is genetically determined; comparing the results of L. spinicrassa (rm = 0.17) with an rm (0.27 daughter females for each cohort female per week) in an established colony of L. shannoni, Cárdenas et al. (1999) indicated that L. shannoni has greater fitness when grown in laboratory conditions than L. spinicrassa. A comparison of the net reproductive rate (Ro = 8.4 daughter females per cohort female), obtained in this work, with L. shannoni (Ro = 23.5) suggests that the population increase of L. spinicrassa under laboratory conditions is very low, and consequently the establishment of the laboratory colonies of this species is very slow and with levels reduced of productivity. It is possible that the stress of colonization reduced the genetic variability that occurs naturally in the field, as it has been demonstrated for several colonies of L. longipalpis (Morrison et al. 1995, Mukhopadhyay et al. 1997) and several mosquito species (Munstermann 1994). The colony environment cannot duplicate conditions of larval nutrition, temperature, humidity, and host availability in the wild and consequently, will strongly affect adult behavior, egg mortality, larval growth period, and adult mortality. Nonetheless, the laboratory data have provided essential baseline information concerning the reproductive potential for L. spinicrassa. For studies of phlebotomine population biology in the field, these observations provide guidelines for approximating the boundaries of expected biological behaviors. ACKNOWLEDGMENTS To Dr María del Pilar Corena, Whitney Laboratory, University of Florida, for her valuable comments and for reading the original manuscript. To the anonymous reviewers whose suggestions greatly improved the manuscript presentation. To Gilberto Torres for the continuous maintenance of the laboratory sand flies colony. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc04127t2.jpg] [oc04127f1.jpg] [oc04127t3.jpg] [oc04127f2.jpg] [oc04127t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}