 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
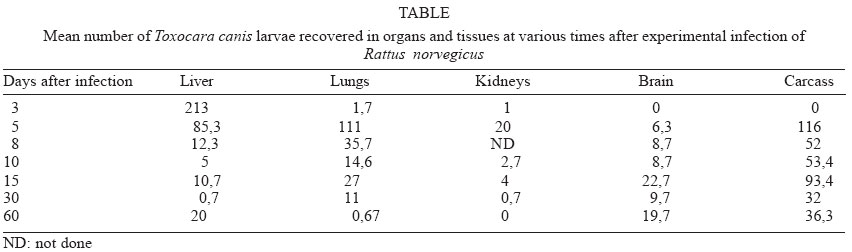
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 6, October, 2004, pp. 627-628 SHORT COMMUNICATION Larval Recovery of Toxocara canis in Organs and Tissues of Experimentally Infected Rattus norvegicus Susana Z Lescano, Maísa L Queiroz, Pedro P Chieffi/*/+ Instituto de Medicina
Tropical, Av. Dr. Enéas de Carvalho Aguiar 500, 2o andar,
05403-000 São Paulo, SP, Brasil Received 22 March
2004 Code Number: oc04130 The aim of this note was to record for the first time the recovery of Toxocara canis larvae from tissues and organs of Rattus norvegicus (Berkenhout, 1769), Wistar strain, until the 60th day after experimental infection. Rats were orally infected with embryonated T. canis eggs, killed on days 3, 5, 8, 10, 15, 30, and 60 after inoculation and larvae were recovered from liver, lungs, kidneys, brain, and carcass after acid digestion, showing a pattern of migration similar of that previously observed in mice. Key words: Toxocara canis - rats - larval recovery - experimental infection Toxocara canis is an intestinal nematode of canids. A wide range of animals, including mice, rabbits, and monkeys act as paratenic hosts. Human infections can occur by accidental ingestion of embryonated eggs of T. canis. The infective larvae hatch, penetrate the intestinal wall, migrate through the soft tissues of the body, and result in either ocular or visceral larva migrans (Glickman & Schantz 1981). Rattus norvegicus, along with other rodents, has been pointed out as usual paratenic host of T. canis (Burren 1972, Chieffi et al. 1981) but there are no reports about the larval distribution of this ascarid in its tissues and organs, other than the central nervous system, after experimental infection. The aim of the present study was to recover the T. canis larvae from tissues and organs of R. norvegicus until the 60th day after experimental infection. Twenty one male R. norvegicus (Wistar strain), with 8 weeks of age, were orally infected with aproximately 500 embryonated T. canis eggs. On days 3, 5, 8, 10, 15, 30, and 60 after inoculation 3 rats were killed by cervical dislocation and larvae were recovered from liver, lungs, kidneys, brain, and carcass after digestion with HCl 0.5% for 24 h at 37oC. Sedimental liquid was poured into a centrifuge tube and centrifuged for 2 min at 1500 rpm, 2 ml of the sediment were collected, thoroughly mixed, and 0.1 ml samples were viewed under the light microscope for larval counts (Wang & Luo 1998). The mean number of larvae recovered from rats is shown in the Table. Larvae from this ascarid were recovered, in higher numbers in the liver (mainly between the 3rd and 5th day after infection), in the lungs (between the 5th and 8th day), in the carcass (since the 5th day), in the brain, since the 5th day after infection and in higher quantity between the 15th and the 60th day after infection. The kidneys showed less larvae between the 3rd and the 15th day after inoculation. Among the different mechanisms described as responsible for the T. canis transmission to canids it has been proposed the possibility of small mammalians to act as paratenic hosts harbouring 3rd stage larvae in their tissues and organs, and transmitting them to canids through the predatory process (Barriga 1988). Most part of the studies carried out to observe rodents as paratenic hosts of this ascarid were done in mice (Koizumi 1984, Guillén Llera et al. 1990, Bardón et al. 1994, Epe et al. 1994). However, R. norvegicus infected with T. canis larvae probably are important as infection source for canids (Chieffi et al. 1981). The migratory route in the mouse includes two phases: a visceral phase, during the 1st week post-infection, when larvae reach the liver and lungs (hepato-pulmonary phase); afterwards, larvae migrate throughout the body and accumulate principally in the carcass and the brain (myo-tropic-neurotropic phase) (Abo-Shehada & Herbert 1985). In our study, we could observe that the higher larvae concentration in the liver, lungs, and kidneys occurred in the 5th day after infection; in the brain and carcass the highest number of larvae was in the 15th day after inoculation, showing a pattern of migration similar of that observed in mice (Bardón et al. 1994, Epe et al. 1994). With this study we can conclude that the recovery of live larvae of T. canis from different organs and tissues of R. norvegicus, up to the 60th day after experimental infection, indicates that this rodent has conditions to act as paratenic host of that ascarid, and may transfer this larvae to a carnivore animal by means of the prey-predator relationship, and may therefore serve as an important factor in the circulation and maintenance of toxocariasis in the biotopes. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc04130t1.jpg] |
| |||||||||

{kind=link}