 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 7, November, 2004, pp. 673-681 Distribution and Schistosoma mansoni Infection of Biomphalaria glabrata in Different Habitats in a Rural Area in the Jequitinhonha Valley, Minas Gerais, Brazil: Environmental and Epidemiological Aspects Helmut Kloos, Liana Kanovaloff Janotti Passos*, Philip LoVerde***, Rodrigo Correa Oliveira**, Andréa Gazzinelli****/+ Department
of Epidemiology and Biostatistics, University of California, San Francisco,
US *Laboratório
de Malacologia **Laboratório de Imunologia e Biologia Molecular, Centro
de Pesquisas René Rachou-Fiocruz, Belo Horizonte, MG, Brasil ***Department
of Microbiology and Immunology, State University of New York, Buffalo, NY,
US ****Escola de Enfermagem, Universidade Federal de Minas Gerais, Av. Alfredo
Balena 190, 30130-100 Belo Horizonte, MG, Brasil Received 16 April
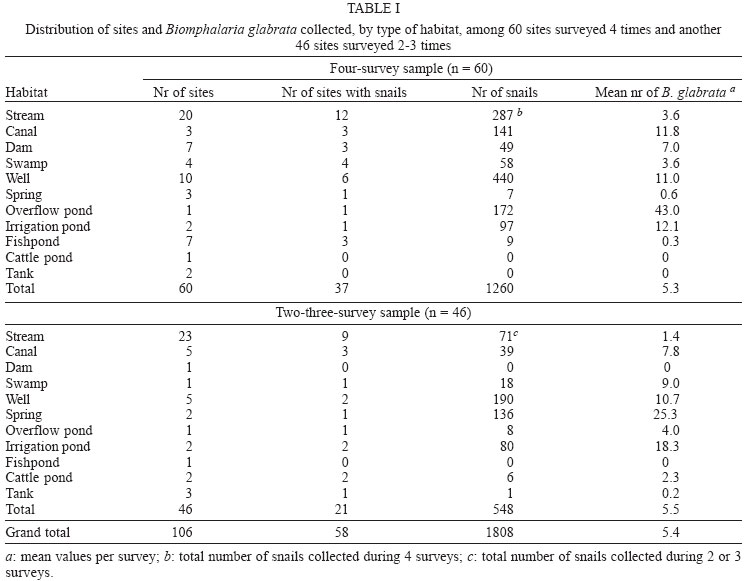
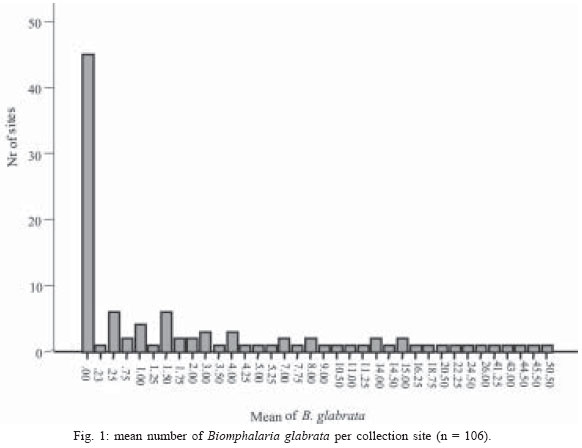
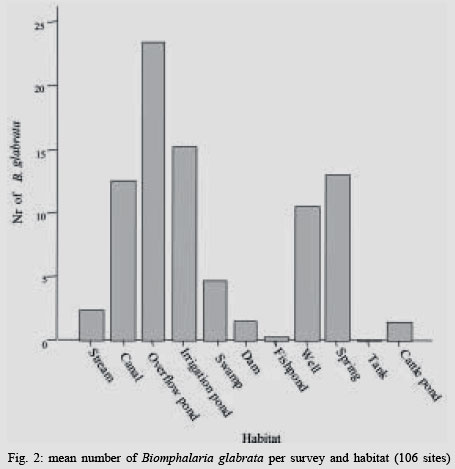
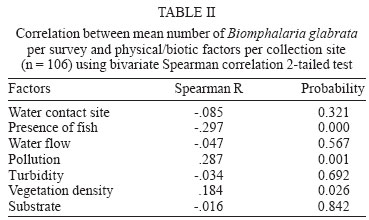
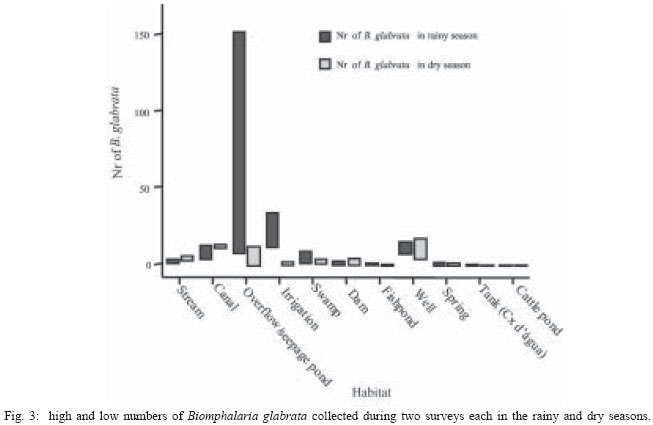
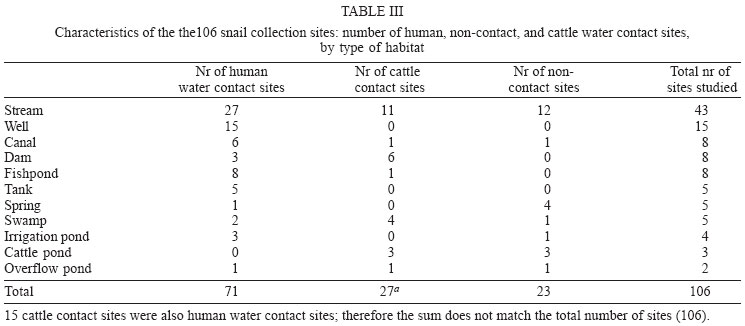
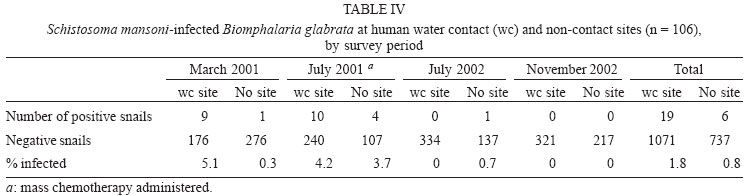
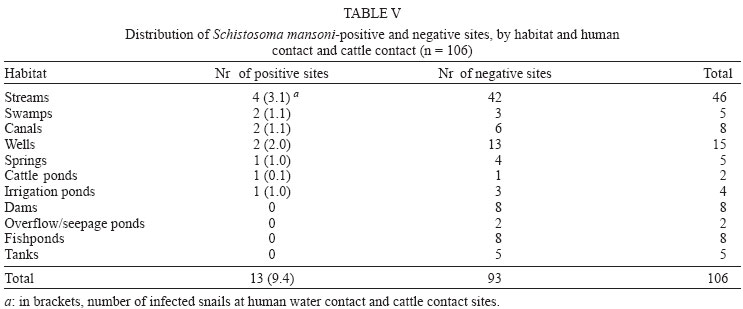
2004 Code number: oc04138 This paper examines the distribution and infection of Biomphalaria glabrata with Schistosoma mansoni in all aquatic snail habitats in a rural area in the state of Minas Gerais, Brazil, in relation to physico/biotic and behavioral factors. Snail and environmental surveys were carried out semi-annually between July 2001 and November 2002 at 106 sites. Collected snails were examined in the laboratory for infection. B. glabrata densities were highest in overflow ponds, irrigation ponds, springs, canals and wells, and lowest in fishponds and water tanks. Snail densities were higher during the hot, rainy season except for streams and canals and were statistically associated with the presence of fish, pollution, and vegetation density. Tilapia fish and an unidentified Diptera larva were found to be predators of B. glabrata but ducks were not. Twenty-four of the 25 infected snails were collected in 2001(1.4% infection rate) and only one in 2002, after mass chemotherapy. The occurrence of B. glabrata in all 11 snail habitats both at and away from water contact sites studied indicates widespread risk of human infection in the study area. In spite of the strong association between B. glabrata and tilapia in fishponds we do not recommend its use in schistosomiasis control for ecological reasons and its relative inefficiency in streams and dams. Key words: Biomphalaria glabrata - snail habitats - environmental factors - tilapia - cattle - Brazil Although there is a large body of knowledge on the distribution of Biomphalaria in different habitats in Brazil, including the state of Minas Gerais and some factors in Schistosoma mansoni transmission (Freitas 1968, Pessoa & Martins 1988, Souza et al. 2001, Carvalho et al. 2003), little is known about the role of tilapia fish, cattle, and physico-biotic factors in snail occurrence and infection, and about the interaction of humans and livestock with snail habitats. Several investigators reported on the involvement of cattle in extra-human transmission of S. mansoni in Brazil and other Latin American and in African countries (Coelho et al. 1982, 1989) and on the predation of Biomphalaria by tilapia and other fish (Andrade & Antunes 1969, Ferguson & Ruiz-Tieben 1971) but most of their findings were derived from laboratory experiments. Pessoa and Martins (1988), who reviewed the literature on the ecology of Biomphalaria species in different aquatic habitats in Brazil, noted the great capacity of species of this genus to adapt to different environmental conditions. In a previous study in a community in northern Minas Gerais we reported the occurrence of B. glabrata and B. straminea in 10 of the 11 types of aquatic habitats studied (Kloos et al. 2001). This paper presents the results of a malacological survey as part of a broader epidemiological study of S. mansoni in the Virgem das Graças study area in the Jequitinhonha Valley, northern Minas Gerais. This large rural area (60 km2) had 131 households and 556 people that were included in the study. Biomphalaria sp. is endemic in most municipalities, in northern Minas Gerais, with B. glabrata the dominant species (Souza et al. 2001, Carvalho et al. 2003) and also the most important vector of S. mansoni in the Americas (Paraense 2001). Fifty-seven percent of the population of Virgem das Graças were infected with S. mansoni in 2001. The major objective of this study was to examine the density and seasonal distribution of Biomphalaria sp. and S. mansoni infections in the study area in relation to physical, and biotic factors and human and livestock water contact behavior. Particular attention was given to the distribution of the intermediate host of S. mansoni in different habitats and the role of predatory fish, particularly tilapia. Whereas some studies pointed to the possibility of fishponds becoming schistosomiasis transmission sites (Editorial 1970, Ferguson & Ruiz-Tieben 1971, Howgate et al. 2002), an increasing number of field studies in Africa (Chimbari et al. 1999) and Brazil (Azevedo 1960, Kloos et al. 2001), indicate that tilapia is an efficient control agent of Biomphalaria in fishponds and lakes. Several pilot control studies involving tilapia and competitors, such as Pomacea lineate, Marisa cornuarietis, and Helisoma duryi, achieved varying results, characterized mostly by ecological upsets after initial snail reductions. MATERIALS AND METHODS All natural and man-made snail habitats and major human and cattle contact sites in the study area were identified during water contact studies and with the help of local informants. Sites were sampled for snails during the rainy season (February/March 2001 and November 2002) and dry season (July 2001 and 2002), at the time when water contact studies were carried out. A standard Fiocruz metal scoop was used and occasionally forceps in rocky and other inaccessible places. Each site was sampled for 5 min per survey by the same person (HK) and all live Biomphalaria sp. larger than 2 mm diameter were collected and identified at the Centro de Pesquisas René Rachou-Fiocruz, Belo Horizonte, and examined for infections twice, at 2-week interval, under artificial light using a 60 Watt bulb for 40-60 min. All snail species were recorded. Water flow was measured midstream by floating styrofoam pellets over a 3 m distance in streams and canals (1 = < 10 cm/s flow; 2 = 10-30 cm/s, and 3 = > 30 cm/s). Samples of substrate were examined visually for texture and composition. Turbidity was visually determined by submerging the metal scoop 5 cm to he depth marked on the handle; if the bright aluminum metal was not visible sites were classified as "highly turbid"; > 5-50 cm (if visible: moderately turbid) and below 50 cm depth: "clear"). Fecal pollution levels were determined based on a combination of smell, visible excretory materials in the water or on the banks, and information obtained from households about drainage of raw sewage into streams. Vegetation density (submerged, floating, and emerging vegetation) was estimated for the specific collection sites in 3 categories (1 = no vegetation, 2 = low to medium density, 3 = high density), and presence of fish was based on a combination of reports by inhabitants of nearby houses, incidental collections with the metal scoop and visual inspection of shallow waters and fish ponds (1 = no fish present, 2 = fish present except tilapia, 3 = tilapia present). Fish incidentally collected were identified with the help of local assistants and returned to the same sites. We observed and interviewed about the presence and range of ducks around households. These measurements and observations were made by the same person (HK) who also carried out the study using the same methodology in the Melquíades study area (Kloos et al. 2001). Using the same pre-tested forms, information was also collected on type of habitat (streams, swamps, canals, wells, springs, cattle ponds, irrigation ponds, dams, overflow ponds, fishponds, and household tanks, or "caixas d'água"), on site location in relation to the nearest study household, whether the sites were used by people, cattle, or both, and the presence of Biomphalaria eggs. Information collected at each site on whether they were used by humans and/or cattle was based on the presence of trampled vegetation, footpaths, the presence of soap stains on rocks, clothe lines, laundry, farm tools, toys, fishing equipment or other water related tools, or animal hoof prints and excrements, as recommended by the World Health Organization (WHO 1979). Bivariate correlation, means, and chi-square tests were applied to examine associations between the distribution of Biomphalaria sp. and environmental factors. RESULTS General distribution - A total of 1808 B. glabrata, the only Biomphalaria species found in the study area, were collected at 106 sites which could be sampled either 4 times (60 sites) or 2-3 times (46 sites). Six sites which were sampled only once or twice during the same season were excluded from the analysis. The 46 sites sampled twice or 3 times were visited at least once each during the rainy and dry season. Absence of families from the home, and flooding, drying up and closure of some sites prevented the sampling of these sites during each survey. Except for springs and overflow ponds, mean numbers of B. glabrata did not differ significantly (p < 0.05) between the 4-survey and 2 to 3-survey samples (Table I). Therefore, most analyses were done on the total data set. Fifty-eight of the 106 sites (54.7%) yielded live B. glabrata and 48 sites (45.3%) were snail-negative. The mean number of B. glabrata found at each snail-positive site per 5 min/1 man of sampling varied from less than 1 to 50, with 10 or more snails collected at 18 sites (Fig. 1). At the same collection sites, 269 Physa snails were also recorded and 4 specimens of a still unidentified snail-predatory larvae species were collected at 3 stream sites. Physa was found in relatively small number at 29 sites, 24 of them populated also by B. glabrata. Distribution by type of habitat - By far the most collection sites were on streams (43), followed by wells (15), canals, dams, fishponds (8 each), swamps, springs and water tanks (5 each), irrigation ponds (4), cattle ponds (3), and stream overflow ponds (2). In Virgem das Graças, streams, canals, springs, wells, and water tanks are all widely used for domestic purposes, canals and streams, swamps and irrigation ponds for agricultural purposes, and many natural waters are often used for playing by children and occasionally for fishing. Preliminary mapping of the spatial distribution of B. glabrata by season using GPS and GIS tools did not reveal significant clustering among sites (A Gazzinelli et al., unpublished observations). The density of B. glabrata per snail habitat was highest in overflow ponds (23.5 snails), followed by irrigation ponds (15.3), springs (13.1), canals (12.6) and wells (10.5), and lowest in fish ponds (0.3) and tanks (0.06). Streams, swamps, dams, and cattle ponds had intermediate densities (Fig. 2). B. glabrata ova were found at 14 sites in 7 habitats (6 stream, 3 well, and 1 canal, swamp, spring, overflow pond, and fishpond without tilapia sites each), indicating the widespread distribution of breeding sites. Physico/biotic factors - In examining the role of physical and biotic factors in B. glabrata distribution, 3 variables - vegetation density, pollution, and presence of fish - were significantly correlated with snail density using a bivariate Spearman correlation test (Table II). Vegetation and presence of fish, vegetation and pollution, substrate and water flow, and turbidity and water flow were all significantly correlated with each other, indicating the interrelationship between these parameters. Seasonal distribution - A significantly larger mean number of B. glabrata was collected in the rainy season (5.8 snails per snail-positive site) than in the dry season (4.6 snails; P < 0.001 in a paired samples t test). However, whereas larger snail numbers were found in standing waters (overflow ponds, irrigation ponds, swamps, springs, and wells) during the rainy season (October-March), densities were higher in streams and canals during the dry season (April-September) (Fig. 3). Of the 37 sites surveyed 4 times which contained B. glabrata at least once, 34 sites were snail-positive in the rainy season and 28 in the dry season (26 of them having snails in both seasons). The significant association between density of vegetation and snail occurrence was influenced by the presence of a large number of B. glabrata snails and ova in water cress (Nasturtium officinale) and of a small number in the dense bulrush (Typha sp., locally known as "taboa") vegetation, swamps, and slow-flowing streams. But large populations of B. glabrata also existed in calcium-rich limestone springs and wells with little or no macro vegetation. Although shell diameter was not recorded for all snails, mean size of B. glabrata was considerably larger in the calcium-rich and low current flow environment of Virgem das Graças than in Melquíades study area. Especially wells and springs contained high calcium levels (as indicated by the substrate and the grey color of the water) and usually large B. glabrata populations in Virgem das Graças, unlike in Melquíades, with a basaltic geology, where this snail was scarce in these habitats (Kloos et al. 2001). Pollution from livestock and sewage was strongly associated with B. gabrata density (P < 0.001; Table II). A form of natural pollution from environmental sources, high ferruginous/alumina levels, tended to reduce snail density but did not eliminate B. glabrata from habitats. None of the 5 fishponds stocked with tilapia yielded B. glabrata. The 3 fishponds without tilapia, located adjacent to a pond stocked with this fish, contained B. glabrata. Similarly, although the supply and drainage canals connected to 2 other fishponds stocked with tilapia contained B. glabrata, the ponds themselves were free of snails. According to local fishermen, tilapia had appeared in recent years and spread in the main Cardoso Stream, which we confirmed when capturing several small tilapia during the snail survey in Cardoso 3 Hamlet. Also "traíra" and "cará" were caught by us at those sites but the relative abundance of these 3 species could not be established. We also collected a small number of B. glabrata at the nearby main water contact site in Cardoso 3, the main source of water for the domestic, recreational, and agricultural needs of the local population. This community had the highest S. mansoni prevalence (90%) in Virgem das Graças study area prior to mass chemotherapy in 2001. Whereas the 3 largest dams (the only other water bodies containing tilapia) were stocked with tilapia for fishing, it is not clear whether the tilapia in Cardoso Stream escaped from fishponds or were introduced by residents for fishing. We collected B. glabrata in one of the dams and found empty shells on the exposed shores of the other 2 dams. Sampling of the dense Typha vegetation was not possible, precluding a full evaluation of the occurrence of B. glabrata in these dams. The age, size, depth, water source (streams, canals, and a spring), and management of fish ponds were not associated with Biomphalaria occurrence. All fishponds were dug in the ground and ranged in size from about 10 to 200 m2 and in depth from about 40 to 80 cm. They were said to be drained annually for a short period for the purpose of harvesting fish and cleaned of silt every 3 to 4 years. Similarly, the practice of regularly feeding tilapia fresh vegetables and food rests in 3 of the 8 ponds was not associated with Biomphalaria occurrence. In addition to tilapia, two other predicted predators of B. glabrata identified in the study were ducks and an unidentified aquatic larva. Only 3 households kept ducks, which foraged in three streams, one dam and one overflow pond. This pond, into which ducks infrequently entered, contained B. glabrata. The confinement of ducks to areas in the immediate vicinity of houses prevented them from feeding at many water contact sites located at greater distance. The unidentified larvae, of the Order Diptera (Prof. Angelo Machado, pers. commun.), found in July and November in streams in Virgem das Graças and in a spring in the Melquíades study area, was observed to be a ferocious predator of B. glabrata in the natural habitat and in captivity, individual specimens killing between 1 and 3 snails per day after their capture (H Kloos, unpublished data). Water contact sites - Sixty-eight of the 106 (64.2%) collection sites, located in 10 of the 11 habitats studied, were human water contact sites and 27 (25.5%) sites in 7 habitats were cattle contact sites (used for drinking, grazing, and crossing). Fifteen cattle contact sites, mostly at streams, dams, and swamps, were also occasionally used by humans (Table III). Cattle were kept away from streams and canals near houses and ponds, in an effort to reduce contamination of the main human water supplies with cattle excrements. None of the B. glabrata density parameters in the aggregate or by survey period were significantly correlated with type of water contact site (either human or cattle sites) in chi-square tests. Snail infection - Twenty-five of the 1808 (1.4%) B. glabrata examined were infected with S. mansoni. All but 1 of the infected snails were found in February/March and July 2001, 1 in July 2002 and none in November 2002. This decrease in infection appears to be due to the administration of mass chemotherapy to all households in the study area in July 2001. Nineteen of the 25 infected snails were collected from human water contact sites and 6 snails from sites not used by humans (Table IV). Thirteen collection sites in 7 habitats had S. mansoni-infected B. glabrata. Infected snails were recovered from several stream, canal, swamp and well sites, and one spring, irrigation pond and cattle pond each. Although no positive snails were found in dams, overflow ponds, fish ponds without tilapia, and water tanks (Table V), they may also provide transmission sites since they all receive water from potentially contaminated streams, canals or springs. Six S. mansoni-infected B. glabrata were collected from 4 sites (one stream, canal, swamp, and cattle pond each) where no human water contact was observed but which were frequently used by cattle for grazing, drinking, and crossing (Table V). DISCUSSION Presence of B. glabrata at collection sites in all 11 natural and man-made habitats studied and of S. mansoni-infected snails at sites in 7 habitats used for domestic, recreational and agricultural purposes indicates widespread transmission in the study area. Year-round occurrence of large numbers of B. glabrata and its adaptation to a wide range of physico/biotic conditions reflect a generally favorable geologic, climatic, and ecological environment in the study area, reported by other investigators in Minas Gerais (Pessoa & Martins 1988, Carvalho et al. 2003). Spatial studies are underway to examine the distribution of B. glabrata in the study area and in different habitats. Some findings of this study corroborate the results of our water contacts studies in Virgem das Graças (Kloos et al. manusc. in preparation), including the widespread use by humans and Biomphalaria of all types of snail habitats both inside communities and in fields and pastures. However, the widespread distribution of B. glabrata and S. mansoni infections in wells and springs, which serve as the major source of domestic water supplies in Virgem das Graças, and differences in the seasonal distribution of snails reveal a different snail ecology and transmission pattern in his study area. The high calcium levels of the predominantly limestone environment of Virgem das Graças appear to promote large B. glabrata populations in wells and springs. Calcium has been associated with high fecundity of B. glabrata (Lévesque et al. 1978) and shell development (Brown 1994). The seasonal distribution of B. glabrata in Virgem das Graças, characterized by generally lower snail densities in standing waters during the dry season but higher densities in streams and canals during that time appears to be determined largely by the reduction of stagnancy through the addition of rainwater in the former and the flushing out of snails by seasonally increased water velocity in the latter. Annual mean maximum temperatures in the nearby town of Araçuai in 2002 were almost 4oC higher during the rainy season (31.9oC) than during the dry season (28.1oC) and higher temperatures during the rainy season were reported from all meteorological stations in Minas Gerais (Instituto Nacional de Meteorologia 2004). This habitat-specific pattern contrasts with the finding by Barbosa et al. (1987) that reproductive rates of B. glabrata were inversely related to temperature. In Virgem das Graças, we noted an inverse relationship between anaerobic conditions (non-filamentous, including toxic algae in warm, shrinking and stagnant water bodies) during the dry season, which are known to reduce snail populations (Brown 1994), and B. glabrata density. Although most streams in the study area swell during heavy rains and may flush out snails, this effect could not be fully evaluated by our semi-annual surveys because of the erratic, irregular rainfall pattern in the Jequitinhonha Valley and the limestone geology of the Virgem das Graças area, which reduces surface runoff after rains. The precipitation effect was more pronounced in areas of basaltic rock and consequently heavier surface runoff elsewhere in Minas Gerais and in Rio de Janeiro (Giovannelli et al. 2001, Kloos et al. 2001). A strong association between pollution and mean density of B. glabrata was noted in spite of dispersed settlement and water contacts in the study area, which tend to disperse and dilute stream water contaminated with sewage and fecal matter at snail collection and water contact sites. The pollution effect was due in part to vegetation density, with which it was significantly correlated (P = 0.003). Similar findings were also reported from an irrigated farm in Rio de Janeiro, where high levels of sewage and quality of water were associated with high B. tenagophila densities (Baptista & Jurberg 1993). Although less numerous in ferrugenous aquatic environments, B. glabrata occurs widely in areas of lateritic soil in Brazil, where it develops brown and black shells. Absence of strong associations between B. glabrata density and water velocity as well as substrate (Table II) is largely due to its successful adaptation of this snail to both standing and flowing waters in the study area. Similarly, both the low-snail density streams and the high-density canals and springs had sandy substrate, and most sites in the other habitats with clay substrate contained a wide size range of snail populations (Fig. 2). Our findings of a strong inverse relationship between the occurrence of tilapia and B. glabrata in fishponds show a combination of ecological effects, particularly predation and type of habitat. They corroborate the field experiments by Azevedo (1960) and Kloos et al. (2001) and the laboratory results obtained by Andrade and Antunes (1969) on the predatory behavior of tilapia against B. glabrata and also indicate their differential impact on snails in different habitats. Both in the Virgem das Graças fishponds and in the 62 fishponds studied in Melquíades, the use of tilapia in extensive traditional and semi-intensive aquaculture was characterized by the raising of fish in relatively small earthen ponds without and with supplementary feed (mostly lettuce and kitchen plant food discards). The increasingly popular intensive aquaculture undertakings in Brazil (especially around cities, which raise large quantities of fish for urban markets by recycling nutrients between pig and/or chicken pens and fish ponds or use commercial pellet feed) or ponds operated for recreational fishing were not observed in Virgem das Graças. In Melquíades, 2 families recycled pig waste in their tilapia ponds. Especially under traditional conditions, tilapia production was associated in both study areas with a "grazed" appearance of macro vegetation in tilapia-stocked ponds, revealing the omnivorous feeding habit of this fish and scarcity of animal food. The relatively long intestinal tract of tilapia species allows them to deal with environmental stress by subsisting on a predominantly herbivorous diet in the absence of the more nourishing and preferred animal food (Hepher 1988, Shiau 2002). Because of their adaptability to a predominantly plant diet tilapia have been reared in the United States for vegetation control (Shepherd & Bromage 1988). In both Melquíades and Virgem das Graças, tilapia was raised in combination with native species such as "traíra", "lambari", "cará", and "bagre" (catfish) and in monoculture, similar to the semi-intensive tilapia farming practices prevailing in most tropical areas. Monoculture assures particularly high yields because of the ability of tilapia to utilize most feeding niches in ponds (Beveridge & Haylor 1998). Although these various studies indicate that tilapia is a potentially useful biocontrol agent in schistosomiasis and vegetation control and a valuable protein source for human diets, its introduction into new habitats and water bodies must be viewed with caution. Tilapia was introduced to Brazil from its African area of origin more than 40 years ago and has caused serious destruction of the indigenous Brazilian fish fauna, especially in the Southeastern region. Absence of significant natural predators in the southeastern provinces has exacerbated the situation in river systems and lakes which tilapia has populated because of its competitive advantages over indigenous species (Cesar AM Estanislau, pers. commun.). Moreover, the presence of Biomphalaria in the main stream in Virgem das Graças, where tilapia appeared several years ago and the high prevalence of S. mansoni in the local population using their waters for domestic, recreational, and agricultural purposes indicates that tilapia is not an efficient biocontrol agent in schistosomiasis control except in animal protein-deficient fishponds. This difference appears to be due to its adaptable omnivorous feeding habits, characterized by a preference for a wide range of animal foods, which are more abundant in streams and other natural water bodies (Stickney 1986). Although relatively little is known about the nutritional requirements and feeding behavior of native Brazilian freshwater fish (Fracalossi 2002), biocontrol studies in schistosomiasis control should give preference to the use of the ecologically more compatible native Brazilian fish species, including Geophagus brasiliensis (Weinzettl & Jurberg 1990), Astronotus ocellatus, and Macropodas opercularis (Consoli et al. 1991). The report by Silveira et al. (1997) that 40% of the fish tanks in a commercial fish farm raising native Brazilian species contained B. straminea cannot be evaluated here as the fish species were not identified. Physa sp., the only other snail species found, does not appear to be effectively competing for habitat with B. glabrata in Virgem das Graças, as indicated by the relatively small number of snails found, its limited distribution, and its coexistence with a larger number of B. glabrata in the same habitats. Ducks also do not play a major role in reducing B. glabrata populations in the study area because of the dispersion of most water contact and snail breeding sites beyond their range, also noted in Melquíades. However, their efficiency in controlling Biomphalaria in other study areas (Kloos et al. 2001), warrants consideration of their use in communities with more focal transmission around residences. The still unidentified Diptera larvas aggressively feeding on B. glabrata resemble behaviorally and cross-anatomically several snail-killing fly species in the Family Sciomiyzidae (Berg 1964) which are widespread in Brazil. Their impact on local Biomphalaria populations is not known. Near-absence of infected B. glabrata one and a half year after mass chemotherapy is consistent with findings of studies in Saint Lucia, where sentinel B. glabrata remained negative for two and a half years after chemotherapy (Barnish 1982). However, this contrasts with a study in Pernambuco, Brazil, which reported no relationship between human infection and snail abundance and infection following chemotherapy (Favre et al. 2002). We conclude that the occurrence of B. glabrata and S. mansoni infections in all habitats studied in Virgem das Graças, both at and away from water contact sites, indicates considerable risk of infection during domestic, agricultural, and recreational water contact activities. The presence of S. mansoni-infected snails at sites used exclusively by cattle or non-contact sites may be due either to transport of cercariae downstream from established human contact sites, incidental human contact away from sites, or contamination by possibly infected cattle, although the role of cattle in transmission still needs to be determined in the study area. Several laboratory studies and field trials in Brazil and in Venezuela found cattle to serve as reservoir hosts of S. mansoni (Coelho et al. 1982, 1989, Modena et al. 1991, 1993, Mott et al. 1995). The limestone geology, climate, and physical and biotic factors operating at the micro-habitat level, may account for the observed distribution of B. glabrata seasonally and among habitats in Virgem das Graças. This study provides preliminary evidence of a differential impact of tilapia predation on B. glabrata distribution in fishponds and streams. In spite of its efficiency in controlling B. glabrata in fishponds, tilapia is not recommended as a biocontrol agent in schistosomiasis control in other habitats in view of the ecological upsets it has caused in the Brazilian fish fauna and its apparent failure to control snails and schistosomiasis transmission in streams. We recommend that controlled field and laboratory experiments focusing on feeding behavior and snail predation using indigenous Brazilian fish species should be carried out instead. ACKNOWLEDGEMENTS To the population of Virgem das Graças for cooperation and assistance, to Ronaldo Gomes Justino, Chris Rowe, and Jennifer Nielsen for assistance with snail collections, to Caroline Patterson with assistance in snail shedding and identification of aquatic plants, and to Professor Paulo MZ Coelho for useful discussions. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc04138t2.jpg] [oc04138f1.jpg] [oc04138t3.jpg] [oc04138f2.jpg] [oc04138t4.jpg] [oc04138t5.jpg] [oc04138f3.jpg] [oc04138t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}