 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 7, November, 2004, pp. 709-715 Effect of the Aedes fluviatilis Saliva on the Development of Plasmodium gallinaceum Infection in Gallus (gallus) domesticus Ana CVM da Rocha/*, Érika M Braga*, Márcio SS Araújo, Bernardo S Franklin, Paulo FP Pimenta/+ Laboratório
de Entomologia Médica, Centro de Pesquisas René Rachou-Fiocruz,
Av. Augusto de Lima 1715, 30190-002 Belo Horizonte, MG, Brasil *Departamento
de Parasitologia, Universidade Federal de Minas Gerais, Belo Horizonte, MG,
Brasil Received 19 April
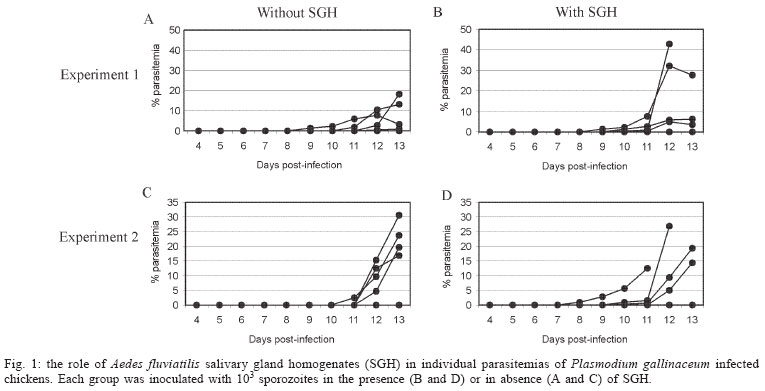
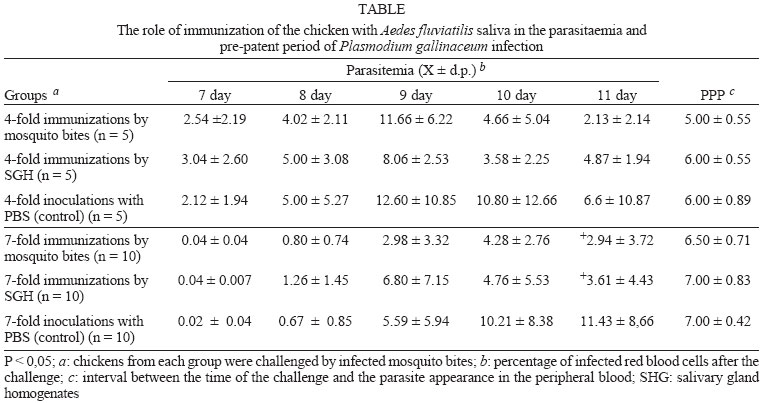
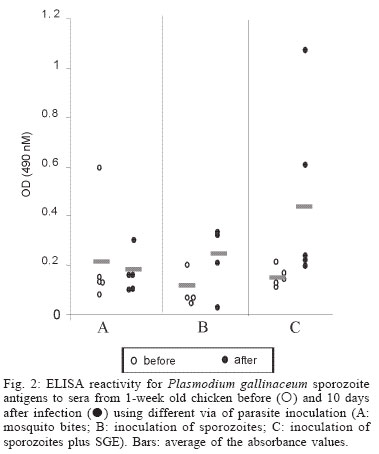
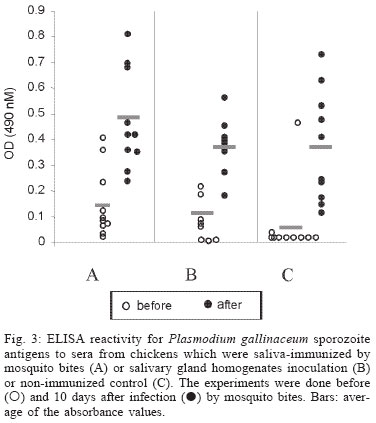
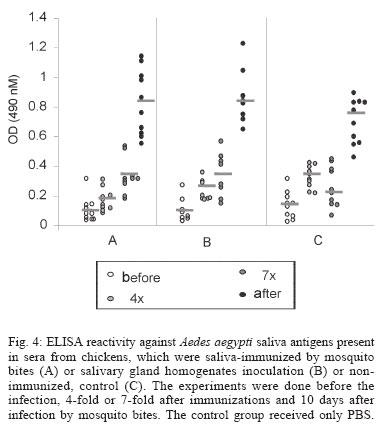
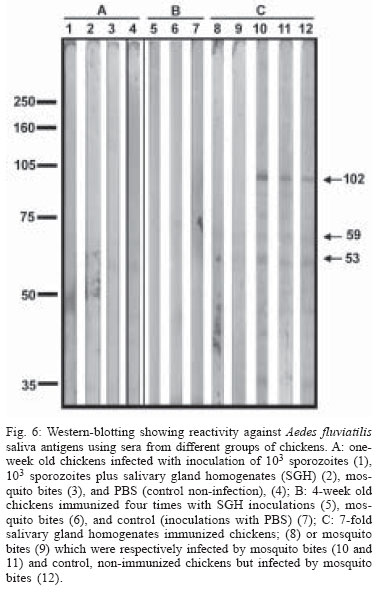
2004 Code Number: oc04144 Effect of Aedes fluviatilis saliva on the development of Plasmodium gallinaceum experimental infection in Gallus (gallus) domesticus was studied in distinct aspects. Chickens subcutaneously infected with sporozoites in the presence of the mosquito salivary gland homogenates (SGH) showed higher levels of parasitaemia when compared to those ones that received only the sporozoites. However, the parasitaemia levels were lower among chickens previously immunized by SGH or non-infected mosquito bites compared to the controls, which did not receive saliva. High levels of anti-saliva antibodies were observed in those immunized chickens. Moreover, 53 and 102 kDa saliva proteins were recognized by sera from immunized chickens. After the sporozoite challenge, the chickens also showed significant levels of anti-sporozoite antibodies. However, the ability to generate anti-sporozoites antibodies was not correlated to the saliva immunization. Our results suggest that mosquito saliva components enhance P. gallinaceum parasite development in naive chickens. However, the prior exposure of chickens to salivary components controls the parasitemia levels in infected individuals. Key words: saliva - avian malaria - mosquitoes - sporozoites - antibodies Saliva components from haematophagous arthropods may modulate the host immune system and would represent an adaptation to evolution related with the blood feeding behavior (Ribeiro et al. 1984, Ribeiro 1987, 1989, James & Rossignol 1991). Several substances in saliva from different vectors such as ticks, phlebotomine, and mosquitoes prevent host homeostasis at the bite site during the blood ingestion. Such components may show different activities such as anti-homeostatic, vasodilator, anti-inflammatory, immune-suppressor, and many others (review in Kamhawi et al. 2000). The effect of saliva substances on the pathogen infectivity to vertebrates was firstly demonstrated by Titus and Ribeiro (1988) in phlebotomine followed by Junes and collaborators (1993) in ticks. Nowadays, the importance of saliva for blood feeding and pathogen infections has been extensively analyzed in several arthropod vectors (review in Gillespie et al. 2000). Therefore, there are few studies about the effect of mosquito saliva on infectivity of Plasmodium parasites. Early studies showed some protection against infection by P. berghei sporozoites when mice were previously immunized with mosquito salivary gland homogenate (Alger et al. 1972, Alger & Harant 1976). In addition, Vaughan et al. (1999) suggested that saliva was one of the factors that could contribute to a more efficient rodent infection when the parasites were injected by mosquito vector bites. In the current study, we evaluate the role of Aedes fluviatilis saliva in the experimental infections of chicken Gallus (gallus) domesticus with P. gallinaceum, the causative agent of avian malaria and its immunogenic potential to control parasitemia. MATERIALS AND METHODS Mosquitoes - Ae. fluviatilis were obtained from a colony established and kept in the Laboratory of Medical Entomology, Centro de Pesquisas René Rachou-Fiocruz in the state of Minas Gerais. The mosquitoes were kept in an acclimated insectary with average temperature between 26-28oC and relative air humidity around 70-80%, in a cycle of 12 h in the dark and 12 h in the light (Consoli & Williams 1978). The mosquitoes were provided with 10% glucose solution and water until the time of the experiments. Mosquito infection - Groups of 4 to 6 day-old female mosquitoes were allowed to feed on the skin of P. gallinaceum infected chickens (parasitaemia levels ranging from 3.5 to 10%). The infection evaluation was carried out 7 to 9 days after the infective blood meal by the presence of ookinetes in dissected midguts, which were dissected in PBS (phosphate buffer solution) and stained with 2% mercury chrome in order to visualize the parasites. The percentage of infected mosquitoes and the number of ookinetes per midgut were recorded through optical microscopic examination. All mosquitoes used to infect chickens were also previously examined for the presence of sporozoites 14 days after the infection (Ozaki et al.1984). Chickens - Newborn G. (gallus) domesticus were acquired from a small commercial farm and maintained in cages. In order to avoid contact of the chickens with any arthropod before and during the experiments, the cages were wrapped with a transparent cotton fabric all the time. Salivary gland homogenate (SGH) - Salivary glands from 4 to 6 day-old female mosquitoes (non-infected mosquitoes) were dissected and transferred to microcentrifuge tubes containing 50 µl cold sterile PBS. Dissected glands were stored at _70oC until use when they were submitted to a successive freezing and unfreezing processes in order to obtain protein material. Total soluble protein concentration in the SGH was determined by Lowry method (Lowry et al. 1951). One salivary gland (a paired structure) dissected from one mosquito was estimated to contain 1.4 µg of proteins. Isolation of sporozoites - The P. gallinaceum sporozoites were obtained according to Ozaki et al. (1984). Briefly, 15 thoraces were dissected from 12-day infected mosquitoes and transferred to 0.5 ml microcentrifuge-tube, containing fiberglass and with perforated bottom, and then placed into another 1.5 ml microcentrifuge-tube. A volume of 50-100 µl of RPMI 1640 culture medium (Sigma, St. Louis, MD, US) supplemented with 10% chicken serum (for experimental infection) or PBS (for antigen preparation) was added to the thoraces. The microcentrifuge-tubes were centrifuged (twice, for 10 min, at 1000 g). The sediment was homogenized and parasites counted into Neubauer's chamber using a phase contrast microscope. The whole process was performed in ice to keep the sporozoite viability. Infection of chickens with P. gallinaceum - One week-old chickens were used to evaluate the infection by P. gallinaceum sporozoites. Groups of five individuals were infected according to the following approaches: (i) naturally infected chickens infected by, bites of 10 infected mosquitoes; (ii) chickens infected by subcutaneous inoculation of 103 sporozoites; (iii) infected chickens by subcutaneous inoculation of 103 sporozoites added with SGH (corresponding to 10 mosquito glands); and (iv) non-infected chickens which were subcutaneous inoculated with RPMI 1640 culture medium. The subcutaneous via was chosen due to the early development of avian malaria into the phagocyte cells in the host skin at the infective bite site (Paraense 1941, 1943, Huff & Coulston 1944, Coulston & Huff 1947). Groups (i) and (iv) were considered as positive and negative controls of the infection. Pre-patent periods (PPP), parasitaemia averages through blood smears stained with Giemsa solution and mortality rates were analyzed and the values compared throughout infection period in the studied groups of chickens. PPP is the period of time between the beginning of the infection and the time that the pathogen is detectable in the peripheral blood. Saliva immunization - The effect of previous exposure to saliva components on the development of infection was analyzed in chickens which received bites from non-infected mosquitoes or which were inoculated by SGH. Groups of 5 or 10 chickens were used according to the following experimental approaches: (i) chickens immunized by bites of 10 non-infected adult mosquitoes; (ii) chickens immunized by subcutaneous inoculations of SGH (corresponding to 10 mosquito glands); and (iii) chickens which received PBS by subcutaneous inoculation. All chickens were one-week old in the beginning of the experiments and they were incubated once a week during a period of 4 or 7 weeks. Sera samples from the chickens were weekly obtained and stored at -20oC until use. One week after the last incubated step, all the chickens received were infected by bites of 10 infected mosquitoes. Detection of anti-sporozoite and anti-SGH IgG - Indirect immunofluorescence assay (IFA) were used to detect anti-sporozoite antibodies and Enzyme linked absorbance (ELISA) were used to detect anti-sporozoite and anti-saliva antibodies in infected or immunized chickens. Briefly, sporozoites were suspended in a concentration of 1x106 parasites/ml and 5 µl were deposited per well on slide. For screening, each well was incubated with 10 µl of sera dilutions (1:20 to 1:5120) in PBS, and then incubated for 20 min with rabbit anti-chicken IgG fluorescein-conjugated (Funed) diluted 1:400 with PBS. The slides were examined by fluorescence microscopy. The ELISA were performed using high binding 96-well microplates (Nunc Maxisorp, Dynatech Denmark) covered with sporozoites homogenate or SGH (20 µg/ml) during 18 h 4°C. Chicken sera were tested in triplicate at 1:80 dilution in PBS-Tween (0.05%) for 2 h at 37°C. The rabbit anti- chicken IgG peroxidase conjugated (Funed) was added at a 1:1000 dilution for 60 min at 37°C followed by addition of peroxidase substrate (OPD-O-phenylenediamine and hydrogen peroxide). Absorbance values were measured at 490 nm. SGH and sporozoite homogenates were also fractionated by electrophoresis SDS-PAGE 12.5% (Laemmli 1970) and then transferred to a nitrocellulose membrane (Amershan Pharmacia HybondTM - C pure). The transference was performed using a 25V constant voltage for 2 h. Part of the nitrocellulose membrane containing the molecular markers was stained with Ponceau S for the visualization of the transferred proteins. The membrane with SGH or sporozoite proteins was incubated for 12 h with chicken sera diluted at 1:40. Afterwards, rabbit anti-chicken IgG peroxidase-conjugated (Funed) was added at a dilution of 1:1000. The antibody reaction was revealed by adding 0.05% 3.3-diaminobenzidin solution containing 0.025%, 4-clhoride 1-naphtol and 0.03% H2 02 (30% v/v). The reaction was interrupted with distillated water after visualizing bands. Statistical analysis - Average parasitemia ands average absorbance were compared using Student's t-test assuming equal variances. Differences in mortality between groups were tested with Chi-square test with Yates' correction. Difference was considered significant when P-value was less than 0.05. RESULTS Effect of the SGH on avian infection - Two distinct experiments were conducted to analyze the effect of the saliva components in the infection of chickens with P. gallinaceum sporozoites (Fig. 1). The parasitemia average among chickens that were infected by sporozoite inoculation in the presence of SGH was higher than those that did not receive SGH (Fig. 1). However, such tendency was not statistically different due to the significant deaths caused by malaria among chickens that were inoculated with parasites plus SGH. The mortality rate of the chickens naturally infected by the mosquito bites was 100% up to the 13 day after infection. Therefore, the mortality rate of chickens infected with sporozoites in the presence of SGH was observed after the 12 day after infection and those infected ones without SGH, only after the 15 day. It is important to point out that approximately 60% of chickens of the group infected by sporozoites inoculation (without SGH) and 40% of chickens of the group infected by sporozoites with SGH, controlled their parasitemia levels at the 18 day after infection (data not shown). The \PPP among chickens naturally infected by mosquito bites was 5 days. Therefore, the PPPs for chickens infected by subcutaneous sporozoite inoculations with or without SGH were around 10 days (data not shown) showing no difference between the two groups. Chicken infection after saliva immunization - Chickens that were immunized 7-fold with saliva by bites or inoculation of SGH showed similar PPP average values after a natural infection by mosquito bites. The average of the parasitaemia levels analyzed at the 9 day after infection of those immunized chicken groups showed to be lower than non-immunized chickens. Statistically significant differences of parasitaemia averages were observed at the 11 day after infection among the groups of immunized chickens and the control group (non-immunized chickens) (Table). The chickens that were immunized four times, either by the mosquito bites or by inoculation of SGH, showed to be more susceptible to the infection than those chickens 7-fold immunized (data not shown). In a follow up of the infection rates during one month, the chickens naturally immunized by mosquito bites presented an average of the mortality rates of 10.5%. Chickens from the SGH-immunized and non-immunized groups showed mortality rates of 12% and 13%, respectively. All the chickens that remained alive in these studied groups appeared to have controlled parasitaemia revealed by negative microscopy examination of blood smears. Anti-sporozoite antibodies in P. gallinaceum infected chickens - Anti-sporozoite antibodies in chicken sera before and after infections were also evaluated. Anti-sporozoite antibodies were not detected by IFA in sera from one-week chickens infected with P. gallinaceum. Fig. 2 shows the individual and the average of sera absorbance values by ELISA verified in a representative experiment. A raise in average values can be observed comparing sera from chickens before and 10 days after infection, independent on the infection via. However, absorbance values average among the experimental groups showed to be relatively low (lower than 0.4). Only two sera from chickens infected by sporozoites with SGH had individual absorbance values higher than 0.6 (Fig. 2). Anti-sporozoite antibodies in saliva-immunized chickens infected with P. gallinaceum - Presence of anti-sporozoite antibodies was not detected among non-infected saliva-immunized chickens. On the other hand, after infection by mosquito bites, all groups of chicken showed anti-sporozoite antibodies including the non-immunized control group. Anti-sporozoite IgG antibodies titles detected by IFA, ranged from 1:40 to 1:1280 in chickens 4-fold immunized reaching a high value (1: 2560) in 7-fold immunized ones. No statistical difference was observed for antibody titles detected by IFA. However, anti-sporozoite antibody frequencies and proportions detected by ELISA showed a statistically significant raise in the average of the absorbance values, which were mainly observed 10 days after infection (Fig. 3). Anti-saliva antibodies in P. gallinaceum infected chickens - Individual values of anti-antibody saliva absorbancies from chickens infected by the mosquito bites or by sporozoite inoculation with or without SGH, showed to be low (lower than 0.4) with no statistically differences among groups (not shown). However, 10 days after infection was observed significant increase in anti-saliva antibody levels among immunized chickens independently of the inoculation via. Such antibody raise were also verified for the control group (Fig. 4). In spite of the number of previous immunizations no statistically significant difference among groups was observed. Recognition of antigenic proteins of P. gallinaceum sporozoite and salivary gland by Western blotting - Sera from chickens infected by P. gallinaceum bites, either from the group of previous saliva-immunized chickens (SGH or mosquito bites) or control group (non-immunized chickens), recognized sporozoite proteins with molecular weights of approximately 64 and 76 kDa (Fig. 5). Which correspond to the P. gallinaceum CS protein. All those sera from chickens of the same groups recognized a protein with a molecular weight of approximately 102 kDa that is presented in SGH (Fig. 6). In addition, proteins of 53 and 59 kDa present in SGH were recognized by the sera from immunized or non-immunized adult chickens infected by the mosquito bites. DISCUSSION The avian malaria infection with P. gallinaceum has been considered a good model for developing studies related with the parasite-host interaction processes. In mental subcutaneous sporozoite inoculation as previously observed (Vaughan et al. 1999). In addition, it is important to consider that sporozoites isolated from salivary glands are considered an heterogeneous population. Moreover, sporozoites need to pass through several biological processes inside the salivary glands in order to reach the salivary duct (Pimenta et al. 1994). Although there are few thousands of sporozoites stocked up in the salivary gland, only a small number is ready to be injected by the mosquito bites (Simonetti 1996). Certainly, not all sporozoites obtained from the salivary gland by the isolation procedure are able to stay alive and develop infection in the skin host. Considering that age-dependent resistance has been demonstrated for pathogens (Kogut et al. 1998, 2002), we evaluated the susceptibility to infection in chickens with different ages. Our results indicate some effect of host age on P. gallinaceum infection: adult chickens showed a prepatent period higher than that observed for young chickens. The young chickens developed the infection in a very quick way with a great number of parasites in the peripheral blood being fatal for several individuals. In opposite, adult chickens appear to be less susceptible to the avian malarial parasite controlling the infection with the absence of deaths. These facts were only observed in infections caused by mosquito bites and not by subcutaneous inoculation of sporozoites, once again, confirming the efficacy of the natural via of infection. It is interesting to note that the presence salivary components in the sporozoite inoculum affect the parasitaemia levels and the mortality rates. An increase in parasitemia levels and in mortality rates was observed when chickens received sporozoites in association with SGH. However, prepatent period, was not affect by the saliva components present in the subcutaneous inoculum. Our results obtained for P. gallinaceum infection, corroborate literature data concerning other pathogens, which evidence the effects of arthropod saliva in parasite-vector interactions (Ribeiro et al. 1985, Titus & Ribeiro 1988, Belkaid et al. 1998, Kamhawi et al. 2000). In our experiments, anti-sporozoite antibodies were detected in chickens infected with P. gallinaceum by mosquito or subcutaneous inoculation. Lower levels of these antibodies were detected in young individuals comparing with the adults. The chickens showed to produce anti-sporozoite antibodies with the same molecular (64 kDa and 76 kDa) weights of the well-known circums-porozoite protein family, which covers the surface of the Plasmodium sporozoite (CS protein). The P. gallinaceum CS protein is involved in parasite interaction with vertebrate and invertebrate cell hosts and elicits a strong humoral response in chicks (Daher & Krettli 1987). In the present study, we demonstrated that adult chickens infected by the mosquito bites produced anti-SGH antibodies. During the blood meal, female mosquitoes deposit saliva into the host skin. Adult chickens sera recognized some SGH proteins including one that correspond to the molecular weight of apirase (64 kDa). Apirases are enzymes that have been demonstrated in the saliva of several insect vectors and are recognized as playing a role in the insect feeding process avoiding the blood platelet aggregation (Ribeiro 1987). Previous work already showed that anti-apirase antibodies in saliva-immunized mice by successive bites of the mosquito Anhopheles stephensi were able to inhibit apirase activity impairing the blood meal screening (Mathews et al. 1996). The parasitaemia levels showed to be lower in immunized groups of chickens than in the control group. Alger and Harant (1976) also reported that immunized mice with mosquito salivary glands were protected against the P. berghei sporozoites challenge. Prior exposure of mice to bites of non-infected sand flies protects against Leishmania major (Kamhawi et al. 2000). However, the sand fly saliva components enhance the cutaneous lesion caused by the parasite. It appears that similar phenomena also occur in saliva-immunized chicken challenged with P. gallinaceum. In conclusion, our results suggest that mosquito saliva components play an important role in the P. gallinaceum infection in chickens. Moreover, saliva also affects the course of P. gallinaceum infection in previously immunized chickens controlling the parasitemia levels. Thus, the role of saliva and its possible use for vaccination against pathogens could be considered. Saliva also can enhance transmission of parasites/pathogens by arthropods. As a result, vaccines that target the arthropod (e.g. salivary immunomodulators) should be considered as one component of multi-subunit vaccines against arthropod-borne pathogens. ACKNOWLEDGMENTS To Denise Nacif Pimenta and Paola Seabra Eiras for correcting the English version. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc04144f5.jpg] [oc04144f4.jpg] [oc04144f2.jpg] [oc04144t1.jpg] [oc04144f1.jpg] [oc04144f6.jpg] [oc04144f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}