 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 7, November, 2004, pp. 717-720 SHORT COMMUNICATION Alteration in the Endogenous Intestinal Flora of Swiss Webster Mice by Experimental Angiostrongylus costaricensis Infection Vandack Nobre, José Carlos Serufo, Omar dos Santos Carvalho*, Cristiane Lafetá Gomes Furtado Mendonça*/***, Simone Gonçalves Santos****, Ester Maria Mota**, Daniel Gomes, Emanuella Braga, Carlos Maurício Figueiredo Antunes*****, Henrique Leonel Lenzi**, José Roberto Lambertucci/+ Serviço
de DIP, Departamento de Clínica Médica, Faculdade de Medicina
*****Departamento de Parasitologia, Instituto de Ciências Biológicas,
Universidade Federal de Minas Gerais, Av. Alfredo Balena 190, sala 3045,
30130-100 Belo Horizonte, MG, Brasil *Laboratório de Helmintoses Intestinais,
Centro de Pesquisas René Rachou-Fiocruz, Belo Horizonte, MG, Brasil
**Departamento de Patologia, IOC-Fiocruz, Rio de Janeiro, RJ, Brasil ***Pontifícia
Universidade Católica de Minas Gerais, Belo Horizonte, MG, Brasil
****Laboratório Micra Biotecnologia, Belo Horizonte, MG, Brasil Received 5 May
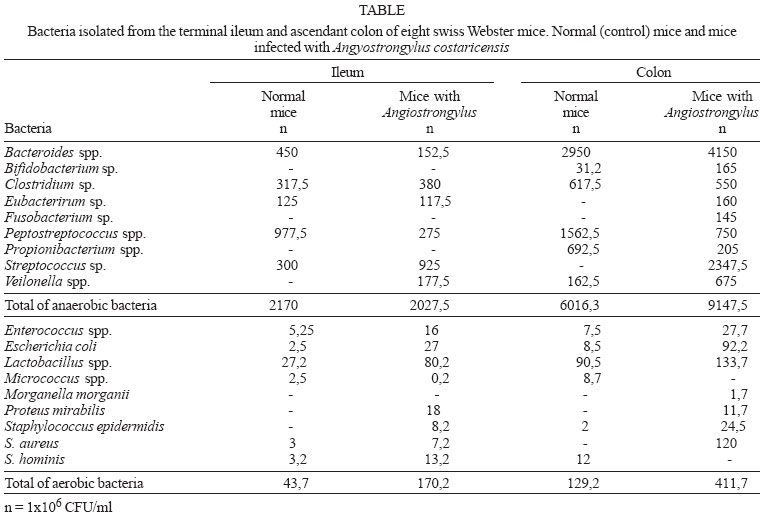
2004 Code Number: oc04145 The association between worm infections and bacterial diseases has only recently been emphasized. This study examined the effect of experimental Angiostrongylus costaricensis infection on endogenous intestinal flora of Swiss Webster mice. Eight mice aging six weeks were selected for this experiment. Four were infected with A. costaricensis and the other four were used as controls. Twenty eight days after the worm infection, all mice in both groups were sacrificed and samples of the contents of the ileum and colon were obtained and cultured for aerobic and anaerobic bacteria. In the mice infected with A. costaricensis there was a significant increase in the number of bacteria of the endogenous intestinal flora, accompanied by a decrease in the number of Peptostreptococcus spp. This alteration in the intestinal flora of mice infected by the nematode may help to understand some bacterial infections described in humans. Key words: Angiostrongylus costaricensis - intestinal flora - Peptostreptococcus Angiostrongylus costaricensis is a rodent nematode, which belongs to the family Metastrongylidae (Morera & Céspedes 1971). Snails from Veronicellidae family act as parasite intermediate host. A. costaricensis is widely distributed in the Americas, being described from the South of the United States of America to the North of Argentina. The worm life cycle in the snails (Mendonça et al. 1999) and rodents (Mota & Lenzi 1995, Mota 2001) has been recently updated. Adult worms have as preferential habitat the blood and lymphatic vessels of the terminal ileum, cecum, and ascendant colon. Abdominal pain and eosinophilia dominate the clinical presentation of abdominal angiostrongyliasis in humans. Diagnosis is based upon the finding of worms and eggs in intestinal specimens obtained during surgery. Enzyme linked immunosorbent assay (ELISA) and polymerase chain reaction (PCR) have also been tested and are available in reference centers (Geiger et al. 2001, Da Silva et al. 2003). Endogenous flora plays an important role in the development of the immunological defense mechanisms of the intestines, as the secretion of IgA, production of epithelial lymphocytes, and the maturation of the MHC molecules (Umesaki & Setoyama 2000). The association between pyogenic abscesses caused by Staphylococcus aureus and worm infection has been described (Lam-bertucci et al. 1990, 2001, Teixeira et al. 2001a,b, Pinilla et al. 2001). Lambertucci et al. (1998) and Rayes et al. (2000) also found an association between tropical pyomyositis and infection by Toxocara canis. At this point, it is worth mentioning the association of Salmonella and other Enterobacteriaceae with Schistosoma mansoni, causing a unique chronic febrile disease, which is cured only after the treatment of the worm. It is also impressive, the recent description of the infection of filarial nematodes, including all of the major pathogenic filariae of humans (Wuchereria bancrofti, Brugia malayi, Onchocerca volvulus) by a bacteria Wolbachia, which lives in the intestine of the worms and seems to play an important role in their development, fertility, and pathogenesis (Taylor & Hoerauf 1999). All these data accentuate the interactions between parasites and different kinds of bacteria as an issue of great importance to improve our understanding of the pathogenesis of some infectious diseases in humans. This study aimed to evaluate the influence of A. costaricensis infection on the intestinal flora of infected mice. Eighteen weeks old male Swiss Webster mice, from the same litter, bred in the animal house of Departamento de Patologia, Instituto Oswaldo Cruz-Fiocruz, Rio de Janeiro, were used for this study. Four mice were orally infected with 8 larvae of A. costaricensis (Crissiumal strain), in accordance with the technique previously described (Mota & Lenzi 1995), and the others were kept as controls (non-infected animals). The following procedures were applied to fecal specimens from all animals, as described below. Twenty-eight days after A. costaricensis infection, mice were submitted to euthanasia by ether inhalation in accordance to animal ethical practice to take intestinal samples. Using aseptic methods, in a biosafety cabinet, the following intestinal segments were isolated: a portion of the terminal ileum and another from the ascendant colon. Each segment measured approximately 3 cm long. After clamping both ends of the segments, a volume of 0.5 ml of sterile saline solution was injected into the lumen using a needle, and after 3 min the contents of the segments were aspirated back. For isolation of strict anaerobes, 0.1 ml of the syringe content was introduced and transported in 0.9 anaerobic transport medium (Ringer's prereduced anaerobically sterilized solution - PRAS). The remainder of the sample was used for isolation of aerobic microorganisms. The time between specimen collection and receipt by the laboratory ranged from 1 to 3 h. An initial dilution of 106 for quantifying bacteria was chosen because the main objective of the study was to evaluate the dominant and subdominant intestinal florae which grow over those levels. The clinical specimens transported in the PRAS solution were introduced into an anaerobic chamber (Forma Scientific Company, Marietta, OH, US) containing an atmosphere of 5% CO2, 10% H2 and 85% N2 and seeded onto the following media: Brucella agar (Inlab, São Paulo, SP, Brazil), Phenylethanol agar (PEA) supplemented with 5 mg/ml of hemine (Inlab), 1 mg/ml of menadione (Inlab) and 5% of sheep blood and Bacteroides-bile-esculin (BBE). The plates were incubated for up to 14 days in an anaerobic chamber at 37ºC. Aerobic bacteria were cultured using MacConkey agar (Difco) and tryptic soy agar (Difco) supplemented with 5% of sheep blood. The plates were incubated aerobically at 37ºC for up to 48 h. Quantitative methods were used for all specimens studied. Identification of microorganisms was performed by biochemical tests and by the semi-automated system Mini-Api (Bio-Merrieux, France). Student's t test was used to compare the means. Due to the exponential characteristics of the bacterial count, data were transformed into logarithm and the geometric means were used for all comparisons. The Table shows the number of the main bacteria (CFU colony formation unit) found in two intestinal segments selected for analysis in this study, separated by genus, in mice infected with A. costaricensis and controls. The isolated bacteria were: Bacteroides spp., Veilonella spp., Clostridium spp., Peptostreptococcus spp., Streptococcus spp., Escherichia coli, Lactobacillus spp., and Enterococcus spp. Some species were found only in the colon of mice, such as, Bifidobacterium spp. and Propionibacterium spp. Interestingly, some bacteria, like Proteus mirabilis and Staphylococcus epidermidis were described only in animals infected with A. costaricensis. In animals infected with A. costaricensis a higher number of aerobic and anaerobic bacteria were observed when the segments of ileum and colon were analyzed together. In the aerobic group there were: E. coli (p = 0.07), Proteus sp. (p = 0.02), Clostridium spp. (p = 0.04). Considering all the anaerobics together, there was a significant increase in the number of bacteria in the group infected with the nematode, except for the Peptostreptococcus spp. which presented a significant lower growth. When anaerobic bacteria in the ileum (p = 0.000019) and colon (p = 0.015) were analyzed independently, a significant difference in the total growth was found in mice infected with A. costaricensis when compared to controls. In the case of Peptostreptococcus spp. exclusion from the analysis, the levels of significance increased both in ileum (p = 0.000002) and colon (p= 0.002). However, no significant comparative differences in these intestinal segments were detected in relation to aerobic bacteria. The present study showed that infection with A. costaricensis interferes with the bacteria normally residing within the lumen of the intestine of Swiss Webster mice, increasing the endogenous bacterial flora and decreasing the number of the anaerobic Peptostreptococcus spp. The panel of bacteria isolated herein did not differ from the normal mice flora described elsewhere (Dewhirst et al. 1999). The worm infection may influence the bacterial growth in the intestine of mice by direct damage to the intestinal mucosa or/and modulation of the immune response of the host as described in helminthic infections (Lambertucci et al. 2001). In fact, severe break of the integrity of the bowel due to ischemic areas, intestinal hemorrhage and perforation were observed in animals infected with A. costaricensis (manuscript in preparation). Direct mechanical lesions caused by adult worms, larvae, eggs, and their secretions may destroy host tissues (e.g., necrosis and granuloma formation) creating favorable sites for adherence of bacteria, facilitating its multiplication and spreading. It appears that immune mechanisms are less important than local tissue lesions, although worm infection can provoke deviation of cytokine profiles and some levels of immunosuppression. The conspicuous behavior of the anaerobic bacterium Peptostreptococcus spp. is demanding explanation. While most bacteria grew at a higher rate in A. costaricensis infected mice, Peptosptreptococcus spp. showed a significant lower growth both in the ileum and colon of these animals. It is well known that this bacterium causes severe infection in humans, and therefore should be the focus of more attention (Murdoch 1998). The translocation of bacteria through the intestinal wall was investigated by Ford (1901). Recently, it has been shown that this phenomenon is an important mechanism of sepsis in humans, ending up in the syndrome of failure of multiple organs. Mucous membrane damage and immunosuppression contribute to such translocation of bacteria (Naaber et al. 2001). It is noteworthy that as part of the normal intestinal flora, Peptostreptococcus spp. has been suggested to have a protective role against infection with pathogenic gram negative bacteria. For instance, Herias et al. (1998) showed that Peptostreptococcus spp. reduces intestinal translocation of E. coli and enhances the production of antibody against the same bacterium in mice. Summarizing, our data show that infection with A. costaricensis alters the intestinal flora of Swiss Webster mice, increasing the growth of normal intestinal bacteria and decreasing the number of the anaerobic Peptostrep-tococcus spp. REFERENCES
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc04145t1.jpg] |
| |||||||||

{kind=link}