 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 99, No. 8, December, 2004, pp. 831-837 Trypanosoma cruzi: Inhibition of a-hydroxyacid Dehydrogenase Isozyme II by N-allyl and N-propyl Oxamates and their Effects on Intact Epimastigotes Miguel A Chena/+++/++++, Silvia Elizondo-Jiménez/+++/++++ , Lorena Rodríguez-Páez/++, Benjamín Nogueda-Torres*/++/+++, Isabel Baeza-Ramírez/++/+++, Carlos Wong-Ramírez/+/++/ +++ Departamento de
Bioquímica *Departamento de Parasitología, Escuela Nacional
de Ciencias Biológicas, Instituto Politécnico Nacional, Apartado
Postal 4-129, Admon. 4, México City, 06401, México Received 5 April
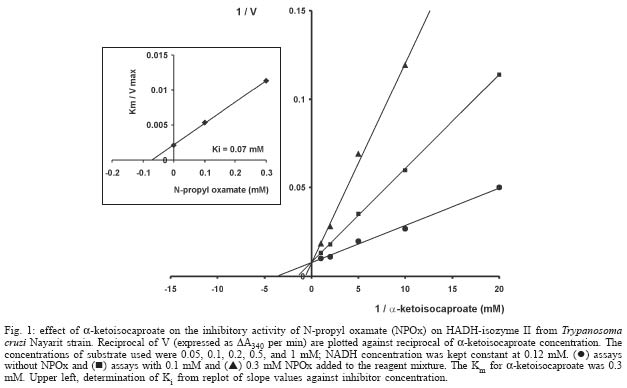
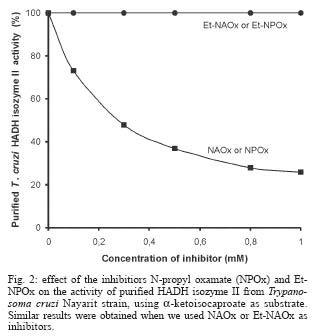
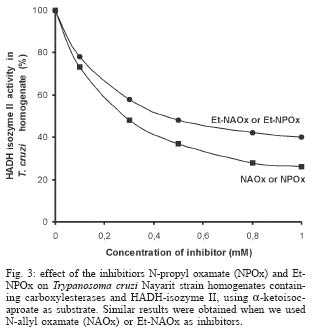
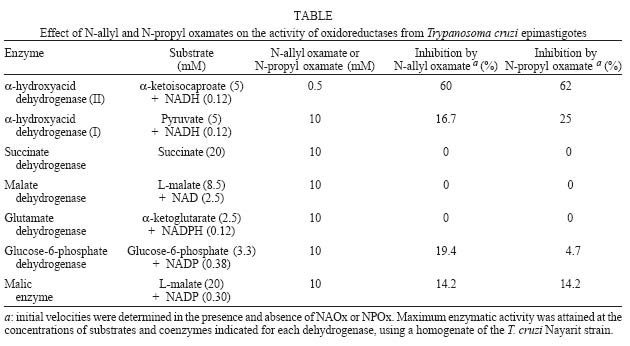
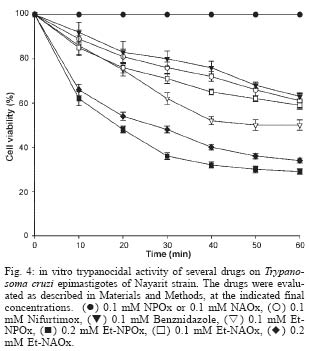
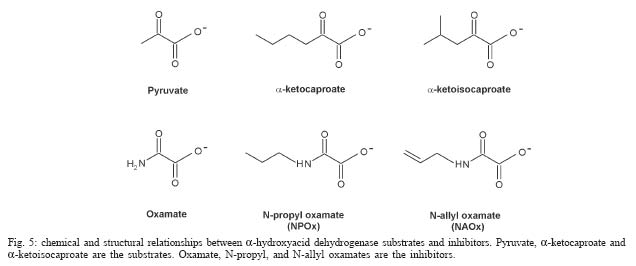
2004 Code number: oc04165 N-allyl (NAOx) and N-propyl (NPOx) oxamates were designed as inhibitors of a-hydroxyacid dehydrogenase (HADH) isozyme II from Trypanosoma cruzi. The kinetic studies showed that NAOx and NPOx were competitive inhibitors of HADH-isozyme II (Ki = 72 µM, IC50 = 0.33 mM and 70 µM, IC50 = 0.32 mM, respectively). The attachment of the allylic and propylic chains to nitrogen of the competitive inhibitor oxamate (Ki = 0.91 mM, IC50 = 4.25 mM), increased 12.6 and 13-folds respectively, the affinity for T. cruzi HADH-isozyme II. NAOx and NPOx were selective inhibitors of HADH-isozyme II, because other T. cruzi dehydrogenases were not inhibited by these substances. Since HADH-isozyme II participates in the energy metabolism of T. cruzi, a trypanocidal effect can be expected with these inhibitors. However, we were not able to detect any trypanocidal activity with these oxamates. When the corresponding ethyl esters of N-allyl (Et-NAOx) and N-propyl (Et-NPOx) oxamates were tested as a possible trypanocidal prodrugs, in comparison with nifurtimox and benznidazole, the expected trypanocidal effects were obtained. Key words: Trypanosoma cruzi - Trypanosoma cruzi a-hydroxyacid dehydrogenase-isozyme II inhibition - N-allyl oxamate - N-propyl oxamate - ethyl N-propyl oxamate - ethyl N-allyl oxamate - a-hydroxyacid dehydrogenase Trypanosoma cruzi, the flagellate parasite that causes Chagas disease, possesses an enzyme similar to the lactate dehydrogenase isozyme-x (LDH-C4) from mammalian spermatozoa (Gerez de Burgos et al. 1976). This T. cruzi enzyme is a NAD-linked oxidoreductase whose catalytic properties resemble those of mouse (Blanco et al. 1976) and rat (Coronel et al. 1983) LDH-C4. The enzyme designated as a-hydroxyacid dehydrogenase (HADH), exhibits two molecular forms (I and II), which have been purified and characterized from the point of view of substrate specificity and kinetic constants (Gerez de Burgos et al. 1978). Isozyme I is responsible for the weak lactate dehydrogenase activity found in T. cruzi extracts (Coronel et al. 1980). On the contrary isozyme II does not show activity against pyruvate and it is active on a broad of linear and branched chain substrates specially a-ketocaproate and a-ketoisocaproate (Coronel et al. 1981). It is interesting that HADH-isozyme II found in T. cruzi, shows a substrate spectrum similar to that of LDH-C4 (Coronel et al. 1981, Montamat et al. 1988), suggesting that this HADH-isozyme II may also accomplish very specific functions in T. cruzi (Montamat et al. 1987). On account of the similarities between HADH-isozyme II and LDH-C4, it has been proposed that these isozymes are functionally homologous (Coronel et al. 1981, Montamat et al. 1987, 1988) and they must be integrated in metabolic pathways supplying energy for the motility of flagellum and survival of the parasite (Coronel et al. 1981, Montamat et al. 1987) or the spermatozoa, respectively (Coronel et al. 1986). HADH-isozyme II is also located in the cytosol and in the matrix of mitochondria (Montamat et al. 1987). On the basis of this dual subcellular distribution, it was proposed that this dehydrogenase, could be also integrated in a shuttle system, transferring reducing equivalents from cytosol to mitochondria, using branched-chain 2-hydroxyacids/2-oxo acids as the redox couple (Coronel et al. 1986, Montamat et al. 1987). Since HADH-isozyme II showed maximum activity with the substrates a-ketocaproate and a-ketoisocaproate (Coronel et al. 1981), we assumed that N-allyl (NAOx) and N-propyl (NPOx) oxamates would fulfill the structural and steric requirements to be competitive and selective inhibitors of T. cruzi HADH-isozyme II, due to the close chemical structure existing between NAOx and NPOx and the substrates a-ketocaproate and a-ketoisocaproate. Thus, we decided to synthesize NAOx and NPOx and to investigate the effect of these substances on the activity of HADH-isozyme II from T. cruzi. This paper presents the inhibitory effect of NAOx and NPOx on HADH-isozyme II, and the effect of these substances on the activity of several oxidoreductases of T. cruzi. In addition, we also tested the possible trypanocidal effect of NAOx and NPOx and that of the corresponding ethyl esters of NAOx (Et-NAOx) and NPOx (Et-NPOx), in epimastigotes of T. cruzi Nayarit strain. The ethyl derivatives are intermediates in the synthesis of the corresponding oxamates. These experiments were run in comparison with nifurtimox and benznidazole, the available drugs for the treatment of Chagas disease. Both drugs are toxic nitroheterocyclic derivatives used as long-term treatment (Teixeira et al. 1994). These drugs cure only a very low percentage of chronic patients (Rodrigues-Coura & de Castro 2002). Natural resistance of T. cruzi to nitro derivatives was suggested as an important factor to explain the low rates of cure detected in chagasic patients (Tsuhako et al. 1991). Since there is presently no satisfactory drug for the treatment of Chagas disease, a very serious endemic affecting millions of people throughout Mexico, Central and South America (WHO 2002), the search for new pharmacological agents is amply justified. MATERIALS AND METHODS Chemicals - NAD, NADP, NADH, NADPH, D-glucose-6-phosphate, pyruvate, oxamate, a-ketocaproate, a-ketoisocaproate, L-malate and L-glutamate all as sodium salts and succinate as the free acid were obtained from Sigma Chemical Co. (St. Louis, MO, US). Other chemicals used were of the highest purity available. Benznidazole (N-benzyl-2-nitro-1-imidazolacetamide) was from Roche-Mexico and nifurtimox (tetrahydro-3-methyl-4-((5-nitro-furfurylidene)amine)-2-methyltetrahydro-1,4-thiazine-4,4 -dioxide) was from Bayer-Mexico. Synthesis of ethyl N-propyl and ethyl N-allyl oxamates - Propylamine or allylamine (0.1 mol) in ethyl ether (50 ml) was added drop wise to an ice cold solution of diethy-loxalate (0.1 mol) dissolved in ethyl ether (100 ml). After being stirred 2 h the reaction mixture was allowed to warm to room temperature and left overnight. A crystalline product separated which was filtered off with suction and the filtrate evaporated under slightly reduced pressure. The residue was then fractionated in vacuum. The Et-NPOx distilled at 80°C at 2 mm, as a colorless oily product. The yield was 85.5%. 1H-NMR (CDCl3) d 0.91 (t, J = 6.0 Hz, 3H), 1.33 (t, J = 5.0 Hz, 3H), 1.57 (qd, J = 7.0 Hz, 2H), 3.20 (q, J = 5.0 Hz, 2H), 4.30 (q, J = 5.0 Hz, 2H). The Et-NAOx distilled at 78°C at 2 mm, as a colorless oily product. The yield was 86.7%. 1H-NMR (CDCl3) d 1.33 (t, J = 7.0 Hz, 3H), 3.96 (t, J = 6.0 Hz, 2H), 4.33 (q, J = 7.0 Hz, 2H), 7.47 (s, 1H). 1H-NMR [Jeol-(JNM-GSX270)]. Synthesis of N-propyl and N-allyl oxamates - Et-NPOx or Et-NAOx (0.05 mol) was shaken with 50 ml of 1 N NaOH for 30 min and extracted with ethyl ether. The aqueous phase was separated and acidified with 2 N HCl. Extraction with ethyl ether and evaporation gave a crude product which, on mixing with light petroleum (range 40-60°C), soon becomes crystalline. The crystals were purified by recrystallization from chloroform. The yield for NPOx was 82%, mp 112-113°C. 1H-NMR (CDCl3) d 0.96 (t, J = 7.0 Hz, 3H), 1.60 (qd, J = 7.0 Hz, 2H), 3.34 (q, J = 7.0 Hz, 2H), 7.25 (s, J = 7.0 Hz, 1H), IR (KBr) 3293, 2981, 1770, 1674, 1556, 1370 cm-1. The yield for NAOx was 84%, mp 97-98°C. 1H-NMR (CDCl3) d 3.98 (t, J = 5.0 Hz, 2H), 5.26 (q, J = 7.0 Hz, 2H), 5.82 (q, J = 7.0 Hz, 1H), 7.41 (broad s, 1H), IR (KBr) 3294, 2970, 1770, 1671, 1552, 1352 cm-1. IR (Perkin Elmer Spectrum GX). Organism - T. cruzi stock Nayarit strain was isolated from Triatoma picturata captured in the state of Nayarit, Mexico. Faeces of infected bugs were inoculated intraperitoneally, into laboratory mice and cardiac blood was cultured subsequently on either enriched biphasic blood-agar medium or the monophasic liquid medium, liver infusion tryptone broth (LIT) supplemented with 10% heat-inactivated fetal calf serum (Chiari et al. 1989). Epi-mastigotes of T. cruzi Nayarit strain were cultivated in the monophasic medium. The parasites were collected from the liquid phase after 7 days of culture. At this stage, the growth is exponential. The liquid phase was centrifuged at 3000 x g for 15 min to obtain the parasites pellet. In vitro evaluation of trypanocidal activity of the drugs - The pellet of parasites was washed 3 times with about 20 parts of 0.15 M sodium phosphate and 0.15 M NaCl buffer pH 7.4. The final pellet of washed parasites was resuspended in 4 parts of the same buffer (1:5) to a final concentration of 1 X 106 organisms/ml. To aliquots (980 µl in eppendorf tubes) of T. cruzi suspension in liquid medium (1 X 106 epimastigotes/ml, final concentration) the drugs dissolved in 20 µl of ethanol, up to 0.1 and 0.2 mM final concentrations were added. Two controls were used: one had ethanol added in the same proportion utilized to dissolve the drugs (20 µl/ml), the other was a parasite suspension without any addition. All samples were incubated at 28°C. The observations and counts were made with a Neubauer hemocytometer after 10, 20, 30, 40, 50 and 60 min incubations (Brener 1962). These experiments were carried out in comparison with nifurtimox and benznidazole. We assumed that immobilized organisms were died because when the immobilized epimastigotes were isolated by centrifugation, washed and reseeded in the same medium LIT we did not observe any growth of these immobilized epimastigotes. On the contrary, when living epimastigotes, from the control experiment were isolated by centrifugation, washed and reseeded in the same medium, extensive proliferation of these parasites was observed. Death of the epimastigotes was also confirmed by the trypan blue dye exclusion method (Urbina et al. 1998). All experiments were run twice at least in triplicate and the results are given as mean ± standard deviation. T. cruzi homogenates - The pellet of parasites was washed 3 times with about 20 parts of 0.15 M sodium phosphate and 0.15 M NaCl buffer pH 7.4. The final pellet of washed parasites was resuspended in 4 parts of the same buffer (1:5) to a final concentration of 1 X 106 organisms/ml. T. cruzi suspension, 1 ml in eppendorf tubes was frozen in liquid nitrogen, and thawed at 37°C. The freeze and thaw operations were repeated 3 times. The disruption of parasites was monitored by microscopic examination of the resulting suspension. The homogenates attained were frozen at -20°C until experiments were performed (usually 24 h later). There was no loss of activity of HADH-isozyme II in the homogenates after storage at -20°C for up to 2 months. Immediately before the enzymatic assays, the suspension was thawed at room temperature and centrifuged at 1200 x g for -20 min at 4°C. The supernatant (0.1 ml properly diluted to obtain a DA340 of 0.05-0.08/min) was used for the analysis of HADH isozymes (I and II). The other dehydrogenases malic enzyme, glucose-6-phosphate, malate, succinate and glutamate dehydrogenases were evaluated in a similar way. Enzyme preparations - HADH-isozyme I and II were purified as described by Coronel et al. (1981), malate dehydrogenase (EC 1.1.1.37) L-malate:NAD oxidoreductase, glutamate dehydrogenase (EC 1.4.1.4) L-glutamate: NADP oxidoreductase (deaminating), malic enzyme (EC 1.1.1.40) L-malate:NADP oxidoreductase (oxaloacetate-decarboxylating), glucose-6-phosphate dehydrogenase (1.1.1.49) D-glucose-6-phosphate:NADP 1-oxidoreductase and succinate dehydrogenase (EC 1.3.99.1) succinate:(acceptor) oxidoreductase, were assayed in the crude extracts of T. cruzi. Enzyme assays - HADH-isozyme II activity was determined using a-ketoisocaproate as substrate (Montamat et al. 1988). The reagent mixture contained, in a final volume of 3 ml: 0.12 mM NADH, 0.1 M sodium phosphate buffer pH 7.4, a-ketoisocaproate as neutral sodium salt (concentrations indicated in results), enzyme preparation, properly diluted with 0.1 M sodium phosphate buffer pH 7.4 in order to obtain an absorbance change at 340 nm of 0.05-0.08 per min with a 5 mM concentration of substrate. Reagent mixtures were incubated at 37°C. Changes in absorption at 340 nm were recorded during a 4 min period. The HADH-isozyme I was determined by the same method using pyruvate as substrate. Succinate dehydrogenase was assayed by the method of DerVartanian and Zeylemaker (1969), glucose-6-phosphate dehydrogenase was assayed by the method of Deutsch (1995), malate dehydrogenase by the technique described by Yoshida (1969), glutamate dehydrogenase by the method used by Juan et al. (1978) and malic enzyme by the procedure used by Cannata et al. (1979). All the enzyme assays were performed at 37°C in order to achieve maximal reaction velocity. However, the conditions selected are also suitable for assays using 25 or 30°C. Determination of Ki values - The enzyme HADH-isozyme II was incubated with the buffer used in the assay, the inhibitors oxamate, NPOx or NAOx and the coenzyme NADH for 10 min at room temperature before starting the reaction by adding the substrate. Values of Ki were calculated from those of Km and V obtained with and without the inhibitor added to the assay mixture. Concentrations of substrates, coenzymes and inhibitors that were used are given in results. RESULTS a-hydroxyacid dehydrogenase isozyme II inhibition - Fig. 1 shows double reciprocal plots of initial velocities at different substrate concentrations (a-ketoisocaproate) and the inhibitory effect of NPOx on HADH-isozyme II. The kinetic studies showed that this substance was really effective inhibitor of this isozyme. The inhibition was shown to be competitive with respect to the substrate a-ketoisocaproate. NAOx was also a competitive inhibitor of this isozyme. The Ki values for oxamate and oxamate derivatives were determined as indicated in Methods and Fig. 1. The Ki value for oxamate was 910 µM, for NAOx was 72 µM and for NPOx was 70 µM. We also determined IC50 values, and there were 4.25 mM, 0.33 mM and 0.32 mM, for oxamate, NAOx and NPOx, respectively. The Km value (0.3 mM) was similar to those reported by others (Montamat et al. 1987). When we used T. cruzi ho-mogenates containing HADH-isozyme II and HADH-isozyme I, the inhibitory effects of oxamate derivatives were the same as those presented for the purified isozymes, using a-ketoisocaproate as substrate for isozyme II and pyruvate for isozyme I (results not shown). We also found that nifurtimox and benznidazole did not inhibit HADH-isozyme II at 5 mM concentration. Effect of N-propyl oxamate and ethyl N-propyl oxamate on the activity of purified HADH isozyme II from T. cruzi - Fig. 2 shows that Et-NPOx it is not an inhibitor of purified HADH isozyme II, whereas increasing concentrations of NPOx (acid) produced increasing inactivation of the enzyme and at 1 mM produced 70% of inhibition. The results with NAOx (acid) were similar to those NPOx and that of Et-NAOx similar to that Et-NPOx. Effect of N-propyl oxamate and ethyl N-propyl oxamate on the activity of HADH from T. cruzi homogenate containing carboxyl esterases - Fig. 3 shows that Et-NPOx was really hydrolyzed by T. cruzi homogenate and the generated derivative (NPOx) really inhibits the HADH present in the homogenate. The inhibition was comparable to that produced by the synthesized NPOx. The results with NAOx (acid) were similar to those NPOx and that of Et-NAOx similar to that of Et-NPOx. Other dehydrogenases activity - We also tested NAOx and NPOx on the activity of other dehydrogenases present in a homogenate of T. cruzi Nayarit strain. At very high concentrations of NAOx and NPOx (10 mM), the malic enzyme, HADH-isozyme I and glucose-6-phosphate dehydrogenase were only slightly inhibited, while glutamate, malate and succinate dehydrogenases were not affected by these substances. On the contrary, crude HADH-isozyme II was inhibited by 60% using 0.5 mM of NAOx, and 62% using 0.5 mM of NPOx, respectively. The Table summarizes the results of the inhibitory action of NAOx and NPOx on the seven studied enzymes, using an homogenate of the T. cruzi Nayarit strain. In vitro trypanocidal activity - Fig. 4 shows the in vitro trypanocidal activity of nifurtimox, benznidazole, NPOx, NAOx, and the Et-NPOx and Et-NAOx against epimastigotes of T. cruzi Nayarit strain. It is observed that at concentration of 0.1 mM, both NPOx and NAOx did not show trypanocidal activity, whereas the corresponding ethyl esters, Et-NPOx and Et-NAOx, showed even better trypanocidal activities than nifurtimox and benznidazole. At a higher concentration, 0.2 mM, both ethyl esters, Et-NPOx and Et-NAOx showed respectively, higher trypanocidal effect. Similar results were obtained with other T. cruzi strains (results not shown). DISCUSSION In this investigation we designed, synthesized, and tested NPOx and NAOx, as well as the ethyl ester of these oxamates on the activity of T. cruzi HADH-isozyme II, because this isozyme has been associated with metabolic pathways supplying energy for the motility and survival of this parasite (Coronel et al. 1981, Montamat et al. 1987). T. cruzi HADH-isozyme I is responsible for the weak LDH activity found in the homogenates of this parasite. The lowest Km was obtained with pyruvate as substrate and does not show activity against a-ketoisocaproate and a-ketocaproate. On the contrary, HADH-isozyme II, the main component of T. cruzi HADH, does not show activity against pyruvate, but it is active on others a-keto and a-hydroxy acids like a-ketoisocaproate and a-ketocaproate, which are the best substrates for HADH-isozyme II, because this isozyme showed maximum activity and lowest Km with this substrates (Coronel et al. 1981). The a-ketoisocaproate and a-ketocaproate are physiological metabolites derived respectively from leucine and isoleucine by transamination, that participate in shuttle systems transferring reducing equivalents from cytosol to mitochondria (Montamat et al. 1987). In addition, HADH-isozyme II it is also an isozyme that participates in the reoxidation of NADH during glycolysis (Coronel et al. 1981). According to this, isozyme II is the isozyme that it is actually integrated in metabolic pathways supplying energy for motility and survival of the parasite (Montamat et al. 1987), and therefore, it has been proposed that inhibitors of this isozyme could reduce the motility and survival of this parasite (Montamat et al. 1982, Blanco et al. 1983, Gerez de Burgos et al. 1984). On account of the close chemical structure of NAOx and NPOx to that of the substrates a-ketocaproate and a-ketoisocaproate (Fig. 5), we assumed that NAOx and NPOx would fulfill the structural and steric requirements to be competitive and selective inhibitors of T. cruzi HADH-isozyme II. The kinetic studies showed that these substances were really effective inhibitors of this isozyme. The inhibitions were shown to be competitive with respect to the substrate a-ketoisocaproate. The kinetic studies also showed that the introduction of the nonpolar allylic and propylic chains in the nitrogen of the oxamate molecule, significantly increased the affinity of NAOx and NPOx for the HADH-isozyme II. Consistent with this were the 12.6 and 13-fold better binding of these oxamates to the active site of T. cruzi HADH-isozyme II. It seems that oxamate, a well known competitive inhibitor of a-hydroxyacid dehydrogenases, will direct NAOx or NPOx molecules against the active site of T. cruzi HADH-isozyme II, because the attachment of the nonpolar allylic or propylic chain to the nitrogen of oxamate will give NAOx or NPOx more affinity and selectivity for the active site of HADH-isozyme II due to the possible hydrophobic interaction of these carbon chain, with a nonpolar region in or near the substrate binding site of this isozyme. Since Baker (1967) was pointed out that the increase in the effectiveness of substrates and inhibitors induced by nonpolar substituents, can be due only to a hydrophobic binding in an enzyme-inhibitor or enzyme-substrate complex. The slightly inhibitory effect of very high concentrations of NAOx or NPOx (10 mM) on glucose-6-phosphate dehydrogenase, HADH-isozyme and malic enzyme, and the lack of any inhibitory activity of NPOx on glutamate, malate and succinate dehydrogenases clearly indicated that NAOx and NPOx were selective inhibitors of T. cruzi HADH-isozyme II, rather than a general inhibitors of dehydrogenases. Our results also suggest the presence of an hydrophobic pocket, similar to that proposed for LDH-C4 (Wong et al. 1997), that can accommodate the nonpolar allylic or propylic group of NAOx or NPOx in the active site of T. cruzi HADH-isozyme II, which might explain the selective inhibition of this isozyme in comparison with the other T. cruzi dehydrogenases and the increasing affinity for those a-keto and a-hydroxyacid substrates containing nonpolar side carbon chains (Coronel et al. 1981). Recently, the glycolytic enzymes have been suggested as a possible target for anti-trypanosomatid drugs design (Bakker et al. 2000, Opperdoes et al. 2001, Verlinde et al. 2001, Lakhdar-Ghazal et al. 2002), because glycolysis provides virtually all the energy for the bloodstream form of trypanosomatids (Michels 1988, Wang 1995, Bakker et al. 2000, Opperdoes et al. 2001). T. cruzi HADH-isozyme II is an important isozyme due to its participation in the energy metabolism of the parasite and therefore, it has been proposed that inhibitors of this enzyme could reduce the motility and survival of this parasite (Montamat et al. 1982, Blanco et al. 1983, Gerez de Burgos et al. 1984, Nogueda et al. 2001). Since in the present investigation we demonstrated that NAOx and NPOx were also selective inhibitors of HADH-isozyme-II from T. cruzi, in this investigation we also tested if these competitive inhibitors also showed trypanocidal activity on this parasite. But when we tested the effect of NAOx and NPOx on cultured epimastigotes we did not detect any trypanocidal activity with these substances. Whereas, with benznidazole and nifurtimox used for comparison, the trypanocidal activity was evident in T. cruzi epimastigotes of the Nayarit strain. However, when we tested the corresponding ethyl esters, Et-NAOx and Et-NPOx, as possible trypanocidal prodrugs we obtained the expected trypanocidal activity, even better than with nifurtimox and benznidazole in this T. cruzi Nayarit strain. At a higher concentration, 0.2 mM, the Et-NAOx and Et-NPOx showed higher trypanocidal effect. Similar results were obtained with other T. cruzi strains (results not shown). Since carboxylesterases activity corresponding to aromatic esterases, and aliphatic or non-specific esterases, has been demonstrated in T. cruzi epimastigotes (Repetto et al. 1983, Aldunate et al. 1987), the increased effectiveness of the Et-NAOx and Et-NPOx to reduce the motility and to kill T. cruzi, most probably results from their better absorption by T. cruzi and its efficient hydrolysis in situ into the active HADH inhibitors (NPOx and NAOx). Using the purified HADH isozyme II from T. cruzi and a T. cruzi homogenate containing the carboxyl esterases and the HADH isozymes we were able to confirm that this was the case, because the Et-NPOx and Et-NAOx were not inhibitors for the purified HADH isozyme II. Whereas when we used the T. cruzi homogenate the esters were rapidly hydrolyzed and the generated acids really inhibited the HADH isozyme II of the T. cruzi extract; these inhibitions were similar to that produced by the synthesized NPOx and NAOx. These experiments also demonstrated that the oxamates did not penetrate on intact trypanosomes, due to the polarity of the carboxylates, whereas the ethyl esters really penetrated inside the parasites where they were hydrolyzed and exerted their pharmacological effect. Additional studies on the action of the Et-NAOx and Et-NPOx on experimental infections of mammalian cell cultures and of animals are necessary to test their activities as possible therapeutic prodrugs in the treatment of trypanosomiasis.
Copyright 2004 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc04165f4.jpg] [oc04165t1.jpg] [oc04165f2.jpg] [oc04165f1.jpg] [oc04165f5.jpg] [oc04165f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}