 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 100, No. 1, February ,2005, pp. 47-49 SHORT COMMUNICATION Genetic polymorphism of the serine rich antigen N-terminal region in Plasmodium falciparum field isolates from Brazil Evelyn Kety Pratt Riccio, Mariano Gustavo Zalis*/++, Helena Cristina Balthazar Guedes**, Dalma Maria Banic, José Maria de Souza***, Wilson Alecrim****, Daniel Camus*****, Cláudio Tadeu Daniel-Ribeiro/++ Maria de Fátima Ferreira-da-Cruz /+/++ Laboratório
de Pesquisas em Malária, Departamento de Imunologia, Centro Colaborador
da OMS para Pesquisa e Treinamento em Imunologia de Doenças Parasitárias,
Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro,
RJ, Brasil *Instituto de Biofísica Carlos Chagas Filho, Universidade
Federal do Rio de Janeiro, Rio de Janeiro, RJ, Brasil **Instituto Nacional
de Controle de Qualidade em Saúde-Fiocruz, Rio de Janeiro, RJ, Brasil
***Instituto Evandro Chagas, Fundação Nacional de Saúde,
Belém, PA, Brasil ****Fundação de Medicina Tropical
do Amazonas, Centro Universitário Nilton Lins, Manaus, AM, Brasil
*****Université de Lille, Laboratoire de Parasitologie-Mycologie,
Faculte de Medecine, Lille, France +Corresponding
author. E-mail: mffcruz@ioc.fiocruz.br Received 9 August
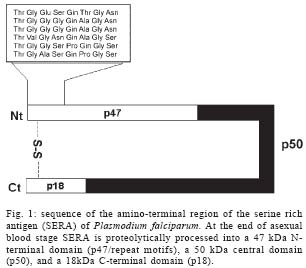
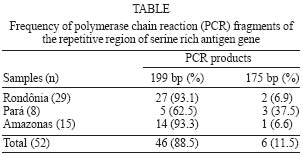
2004 Code number: oc05009 In this work we investigated the frequency of polymorphism in exon II of the gene encoding most of the amino-terminal region of the serine rich antigen (SERA) in Plasmodium falciparum field samples. The blood samples were colleted from P. falciparum infected individuals in three areas of the Brazilian Amazon. Two fragments have been characterized by polymerase chain reaction: one of 175 bp corresponding to the repeat region with 5 octamer units and one other of 199 bp related to the 6 repeat octamer units of SERA protein. The 199 bp fragment was the predominant one in all the studied areas. The higher frequency of this fragment has not been described before and could be explained by an immunological selection of the plasmodial population in the infected individuals under study. Since repeat motifs in the amino-terminal region of SERA contain epitopes recognized by parasite-inhibitor antibodies, data reported here suggest that the analysis of the polymorphism of P. falciparum isolates in different geographical areas is a preliminary stage before the final drawing of an universal vaccine against malaria can be reached. Key words: malaria - Plasmodium falciparum - serine rich antigen - polymorphism Malaria is still nowadays one of the most important problems of public health in endemic areas. As a result and because of the emergence of resistance of both the parasite and the mosquito vector to drugs and insecticides, respectively, a malaria vaccine is one of the most powerful potential tools to be added to those classically used to control malaria transmission. Consequently, different antigens expressed during the asexual cycle of the malaria parasite and their encoding genes have been characterized in the last years (Ferreira et al. 1998, Sallenave-Sales et al. 2000, Magesa et al. 2002). The gene codifying to the serine rich antigen (SERA), a protein also known as p126 or serine-rich protein (SERP), which is located in the parasitophorous vacuole of trophozoites and schizonts, has been also target of interest to several groups. SERA ranks as a candidate antigen for inclusion as a subunit in a polyantigen malaria vaccine because: (i) specific monoclonal and polyclonal antibodies against SERA can inhibit the in vitro growth of the parasite (Chulay et al. 1987, Bzik et al. 1988, Knapp 1989); (ii) immunization with SERA can induce partial protection against parasite challenge in Saimiri and Aotus monkeys (Perrin et al. 1984, Delplace et al. 1985, Inselburg et al. 1991, Enders et al. 1992, Knapp et al. 1992); and iii) a positive association between infection induced antibody response and the degree of protective immunity has been reported (Banic et al. 1998, Okech et al. 2001). The SERA gene, firstly isolated from the genomic DNA of FCR3 strain, is localized in the chromosome 2 of the P. falciparum genome (Biggs et al. 1989). Although SERA showed a quite conserved sequence in P. falciparum isolates from different geographical origins including Asia, Africa, and South America (Bhatia et al. 1987, Delplace et al. 1988), two regions of polymorphism have been observed in different P. falciparum laboratory samples. The registered polymorphism comprises events of deletions/insertions in two repetitive regions of the protein: one in the polyserine region/serine repeats (SR) and one other in the amino-terminal region/octamer repeats (OR) that could comprise 5 instead of 6 octamer units (Fig. 1) (Bizik et al. 1988, Knapp et al. 1989, Morimatsu et al. 1997, Safitri et al. 2003). Basically, the number of OR, containing 8 amino acids each one, characterizes the polymorphism: the allele I, with 5 repeats, and the allele II, with 6 repeats (Li et al. 1989). This later region was showed to be very immunogenic (Banic et al. 1998) and is involved in the induction of protective immunity in non-human primates (Inselburg et al. 1993a,b, Suzue et al. 1997). Considering the potential inclusion of SERA in a malaria vaccine together to the fact that repeat motifs in the amino-terminal region of SERA contain epitopes recognized by parasite-inhibitor antibodies (Fox et al. 2002), the study of the genetic diversity of this protein is mandatory, since sequence variation in exon II may represent one of the parasite's immune-evasion strategies. By combining previously published sequences, it has been observed that FCR3 type alleles predominated in P. falciparum field isolates from Indonesia, Brazil, and Solomon Islands but have not been found in Myanmar and Africa (Safitri et al. 2003). Here we have investigated the frequency of polymorphism in exon II of SERA gene, which encodes most of the amino-terminal region of the antigen in P. falciparum field samples. The blood samples were colleted from P. falciparum infected individuals in three areas of the Brazilian Amazon: Porto Velho, at the state of Rondônia; Belém and Marabá, at the state of Pará; and Manaus, at the state of Amazonas. The patients were assisted at the Centro de Medicina Tropical de Rondônia in Porto Velho (n = 29), at the Instituto Evandro Chagas of the Secretaria de Vigilância em Saúde in Pará (n = 8), and at the Fundação de Medicina Tropical do Amazonas in Manaus (n = 15). Patients were invited to participate through the "Term of Post-informed Consent", informed of the objectives, and the role of their participation in the study and written consents were obtained. After submitting to each patient a questionnaire, which included personal history and epidemiological data, a venous blood sample (5 ml) was collected from each individual using vacutainer tubes containing EDTA. After centrifugation, packed red blood cells (RBC) were separated for DNA extraction and PCR analysis, and all the samples were cryopreserved in glycerolyte (w/v) in duplicate and stored in liquid nitrogen tank, until use. By using PCR it was possible to characterize two fragments (Fig. 2): one of 175 bp corresponding to the repeat region with 5 octamer units (allele I) and one other of 199 bp corresponding to the 6 repetitive octamer units (allele II) of SERA protein. As shown in the Table, the 199 bp fragment was the predominant one in all the studied areas. The higher frequency of this fragment could be explained by an immunological selection of the plasmodial population in the infected individuals under study (Daubersies et al. 1994). We should emphasize that mixed infections have not been observed in any of the isolates studied. In conclusion, if the here reported sequence polymorphism affects the immune recognition of SERA, the present results indicate that the analysis of the polymorphism of P. falciparum isolates is a fundamental stage before the final drawing of an universal vaccine against malaria can be reached. ACKNOWLEDGMENTS To those who kindly provided blood samples for this study. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05009t1.jpg] [oc05009f2.jpg] [oc05009f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}