 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
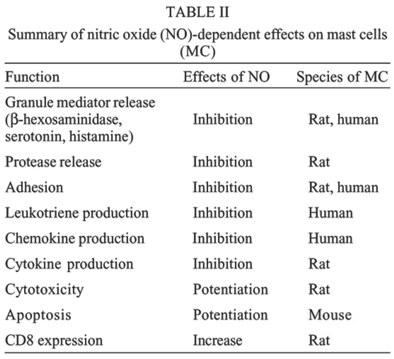
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol.100, Suppl. 1, March, 2005, pp. 11-14 Nitric oxide: a major determinant of mast cell phenotype and function SD McCauley, M Gilchrist*, AD Befus/+ Department of Medicine, Pulmonary Research Group, University of Alberta, 567 Heritage Medical Research Center, Edmonton, Alberta, Canada, T6G 2S2 *Institute for Systems Biology, Seattle, WA, US +Corresponding author. E-mail: Dean.Befus@ualberta.ca Received 8 November 2004 Code number: oc05020 Mast cells (MC) are important in the numerous physiological processes of homeostasis and disease. Most notably, MC are critical effectors in the development and exacerbation of allergic disorders. Nitric oxide (NO) is a diatomic radical produced by nitric oxide synthase (NOS), and has pluripotent cell signaling and cytotoxic properties. NO can influence many MC functions. Recent evidence shows the source of this NO can be from the mast cell itself. Governing the production of this endogenous NO, through alterations in the expression of tetrahydrobiopterin (BH4), a NOS cofactor, has stabilizing effects on MC degranulation. Furthermore, NO regulates the synthesis and secretion of de novo generated mediators, including leukotrienes and chemokines. These novel observations add to the growing body of knowledge surrounding the role of NO in the MC. Key words: nitric oxide - mast cell - tetrahydrobiopterin - leukotriene - degranulation Nitric oxide (NO) is a potent mediator with diverse roles in regulating cellular functions, including ni-trosylation of proteins involved in signaling pathways (Alderton et al. 2001, Stamler et al. 2001). NO is produced by the oxidation of the amino acid L-arginine by the enzyme nitric oxide synthase (NOS). This family of enzymes are generally classified as constitutive, calcium dependent (neuronal NOS, nNOS, NOS1 and endothelial NOS, eNOS, NOS3) or inducible, calcium independent (inducible NOS, iNOS, NOS2) (Alderton et al. 2001). The constitutive isoforms release relatively low levels of NO, while iNOS is induced by inflammatory conditions and releases larger amounts of NO. Mast cells (MC) arise from hematopoeitic precursor cells in the bone marrow and mature and reside in tissues (Kulka & Befus 2003). They produce, store, and secrete a large spectrum of biologically active mediators and have roles in many physiological processes such as tissue remodeling, wound healing, angiogenesis, and innate immunity (Wedenmeyer et al. 2000). Due to their bioactive armamentarium and location within tissues, MC are also major players in numerous disease states, of which TH2-mediated inflammatory conditions like allergy and asthma are best known (Brightling et al. 2003). Our lab and others have shown that NO regulates MC function (Bissonnette et al. 1991, Brooks et al. 1999, Coleman 2002, Gilchrist et al. 2003, 2004). In addition, we have demonstrated the expression of NOS isoforms and production of NO by various MC populations including rat peritoneal MC (PMC), human skin MC (HSMC) and human mast cell lines (HMC-1 and LAD-2) (Gilchrist et al. 2002, 2004). Furthermore, we investigated the role endogenous NO has on various MC functions including cytotoxicity, adhesion, early mediator release, and leukotriene production (Bissonnette et al. 1991, Forsythe & Befus 2003, Gilchrist et al. 2004). NOS require various co-factors to function, including calmodulin (CaM), FAD, FMN, and iron protoporphyrin IX (haem) (Alderton et al. 2001). Additionally, tetra-hydrobiopterin (BH4) is required for NOS activity and is produced by the enzyme GTP-cyclohydrolase I (CHI) (Gilchrist et al. 2003). Regulation of constitutive NOS function in human MC appears to play an important role in decreasing the production and secretion of leukotrienes (LT) in a cGMP independent way (Gilchrist et al. 2004). Furthermore, NO is able to inhibit protease release (Kanwar et al. 1994) and chemokine (M Gilchrist, unpublished observations) and cytokine (Coleman 2002) production by MC. NO can regulate autocrine apoptotic signals (Kitajima et al. 1994), and alter surface molecule expression, namely CD8 (Nohara et al. 2001). It is apparent that NO is important in both allergic and non-allergic functions of MC. This review will summarize recent findings from our lab and others describing NO-dependent alterations in MC function. MATERIALS AND METHODS MC studied - Adult male Sprague-Dawley rats were obtained from Charles River Canada Inc. (Quebec, Canada). PMC were obtained by peritoneal lavage and Percoll purification. Rat cultured MC (RCMC), a gift from Drs A Froese and B. Chan, University of Manitoba were cultured as described previously (Chan et al. 1990). Mouse bone marrow cultured MC (BMMC) were grown using the protocol of Vliagoftis (2002). The growth factor independent human MC line (HMC-1) were cultured in Iscove's medium with 10% fetal bovine serum (FBS), 2mM glutamine, 40U/ml penicillin/streptomycin and 1.0 mM thioglycerol. The growth-factor dependent human MC line LAD-2 (obtained from Dr AS Kirshenbaum and DD Metcalfe, NIH, Bethesda, MD) (Kirshenbaum et al. 2003), was cultured in StemPro-34 medium supplemented with 100ng/ml recombinant stem cell factor (rhSCF). Both human cell lines were maintained at 37oC in a humidified incubator with 5% CO2. Human skin MC (HSMC) were isolated and enriched (³ 93% purity; Gilchrist et al. 2004). Measurement of NO - (i) DAF-FM, a cell permeable fluorescent dye, was used to assay NO production (Suzuki et al. 2003) in PMC with live cell microscopy using a Zeiss confocal laser scanning microscope (LSM510); (ii) NO2- levels in culture supernatants were measured by the Griess reaction (Gilchrist et al. 2002). Reverse transcription-PCR - Total RNA was isolated from PMC and human MC populations as previously described (Gilchrist et al. 1997). RNA was converted to DNA by the reverse transcription reaction (M.MLv reverse transcriptase, Invitrogen, Burlington, ON). PCR amplification was performed on a PTC-100 Thermal Cycler (MJ Research, Boston, MA). All primers were designed to be intron spanning based on published sequence data. All products were sequenced and confirmed (Gilchrist et al. 2004). ELISA for cysteinyl leukotrienes - HMC-1 and LAD-2 were stimulated with calcium ionophore (A23187) or anti-IgE, respectively. Culture supernatants were collected and cysteinyl leukotriene (LTC4, D4, E4, C5, D5, E5) production was quantified using EIA (Cayman Chemical) according to manufacturer's protocols. RESULTS Expression of NOS in MC - Confocal laser microscopy, immunoblotting, and RT-PCR studies of both rodent and human MC populations have identified expression patterns of the three NOS isoforms (Table I). Regulation of GTP-CHI expression modulates BH4 production, NO production, and MC function - Exposure of PMC to IFN-g (200 U/ml) for 18 h increases GTP-CHI mRNA, protein, and enzymatic activity and decreases GTP-CHI feedback regulatory protein (GFRP), a GTP-CHI inhibitor, production (Gilchrist et al. 2003). This increase in BH4 production correlates with an increase in NO production, measured by nitrate production, and this effect can be mimicked by addition of an exogenous source of BH4, sepiapterin. Increased DAF fluorescence, indicating endogenous NO production, is seen in the majority of sensitized PMC treated with IFN-g. NO producing MC are more refractory to antigen stimulated degranulation signals than non-NO producing MC (Gilchrist et al. 2003). Expression and location of NOS in human MC affects leukotriene production - Confocal analysis was used to determine the location of constitutive NOS (cNOS) expression in HMC-1 and LAD-2 cells. We were unable to detect nNOS in LAD-2 (Table I), whereas HMC-1 showed diffuse cytoplasmic staining for nNOS. eNOS was identified in both LAD-2 and HMC-1, predominantly nuclear in location, and was confirmed through immunoblot analysis of subcellular fractions. LT are produced by an enzymatic cascade which involves 5-lipoxygenase (5-LO), an enzyme known to be located in the nucleus of a number of different cell types (Peters-Golden & Brock 2001). NO and 5-LO have been shown to interact previously in other cell types (Coffey et al. 2002). 5-LO translocated to the nucleus of HMC-1 by 15 min after A23187 stimulation. This accompanies a calcium dependent phosphorylation of eNOS, known to increase its activity. Nuclear co-localization of these two enzymes was seen using confocal microscopy after A23187 treatment (HMC-1) and IgE-crosslinking (LAD-2). These same stimuli resulted in production and release of LT from both cell types. Pretreatment of the cells with a NO donor (SNOG) resulted in a dose-dependent decrease of LT production. Conversely, addition of a NOS inhibitor, L-NAME, resulted in a dose-dependent increase in LT production. The cGMP analogue, 8-bromo-cGMP, had no effect on LT release from either cell type. Further NO-mediated changes in MC function - In addition to the recent observations mentioned above, NO plays an important role in a number of other MC-mediated events. A summary is provided in Table II. Adhesion: activated MC are able to bind to extracellular matrix proteins, including fibronectin (FN). We have shown that treatment of the rat MC line RBL-2H3 with IFN-g and HMC-1 with various NO donors abrogates the ability of both cell types to adhere to FN (Wills 1999, Forsythe & Befus 2003). In the latter case, the effects were mediated through the nitrosylation of the cysteine protease calpain in a cGMP-independent manner (Forsythe & Befus 2003). Granule mediator release: in addition to our observations, Brooks et al. (1999) have shown that pre-incubation of PMC with the NOS inhibitor L-NAME causes a significant increase in histamine release when stimulated by various chemical and antigenic stimuli. Furthermore, recent observations in our lab have demonstrated a NO-dependent regulation of b-hexosaminidase release (a marker of degranulation) from LAD-2 cells (SD McCauley, unpublished observations). Finally, Deschoolmeester et al. (1999) showed that IFN-g and NO-donor treatment of a mixed mouse peritoneal cell population inhibited MC serotonin release in a cGMP-independent manner. Protease release: in assessing the involvement of MC on intestinal epithelial permeability, Kanwar et al. (1994) determined that increased permeability was associated with an increase in rat MC protease II (RMCPII). L-NAME greatly enhanced this RMCPII release, and this effect was attenuated by the addition of MC stabilizers, suggesting NO regulates this process. Chemokine and cytokine production: recent observations in our lab have suggested a role for endogenous NO in regulating production of chemokines in human MC. HMC-1 treated with IFN-g showed decreased mRNA expression and secretion of both MCP-1 and I-309, and these effects were eliminated with pre-treatment with L-NAME (Gilchrist M, unpublished observations). Moreover, Coleman (2002) has reported evidence NO-donors inhibit antigen-induced mRNA expression of IL-4, IL-6, and TNF from RBL-2H3 cells. Apoptosis/cytotoxicity: NO is able to increase MC-mediated cytotoxicity, as PMC killing of a TNF sensitive target cell (WEHI-164) was inhibited by the addition of L-NAME and was enhanced by either addition of NO-donors or L-arginine (Bissonnette et al. 1991). Additionally, mouse MC undergoing apoptosis are dependent on endogenous NO signals, as NOS inhibitors prevented apoptosis from occurring (Kitajima et al. 1994). CD8 expression: our lab has previously reported the expression of CD8, the ligand/receptor for MHC class I, on the surface of PMC (Lin et al. 1998). Further studies have demonstrated NO increases the expression of CD8, after the ligation of existing CD8 with a specific antibody, in a cGMP-dependent manner (Nohara et al. 2001). DISCUSSION Several years ago we and others established that NO modulates MC function (Salvemini et al. 1991, Bissonnette et al. 1991), though the precise source of this NO has been controversial. Using various methods, our lab has demonstrated that MC themselves express NOS mRNA and protein and have accompanying production of NO (Gilchrist et al. 2002, 2003, 2004). Others have clearly shown that macrophage-derived NO can be important (Eastmond et al. 1997, Coleman 2002). There appears to be significant heterogeneity in NOS expression among MC populations (Table I). Rat MC are capable of expressing iNOS, whereas HSMC and human MC lines primarily express one or both of the constitutive NOS isoforms. To date, we have been unable to detect iNOS in human MC. This suggests there may be important differences between rat and human MC in NO production and NO-mediated regulation of MC function. Low NO levels produced by cNOS are sufficient to alter MC function (Gilchrist et al. 2004). Endogenous production of NO affects function of MC. PMC producing iNOS and NO following exposure to IFN-g are more resistant to stimuli that produce degranulation than are those cells not producing NO (Gilchrist et al. 2003). When stimulated with A23187 (HMC-1) and IgE-crosslinking (LAD-2) human MC lines produce LT. NO produced by these cells directly influences this LT synthesis and secretion (Table II). Because these effects are independent of cGMP, and given that 5-LO and eNOS co-localize, direct nitrosylation of 5-LO, shown previously to occur in macrophages (Coffee et al. 2002), may be responsible (Gilchrist et al. 2004). In addition to these novel observations, NO is crucial to numerous other MC functions, including degranulation, adhesion, and protease release, as well as production and secretion of chemokines and cytokines (Table II). Combined, these data suggest NO mediated regulatory and signaling events are critical in the development of allergic responses driven by MC. Finally, it is apparent NO also plays a role in modifying functions in MC not associated with allergic disease, as MC-mediated cytotoxicity, apoptosis, and surface protein expression are closely linked to NO production (Table II). Interestingly, not all of NO effects are cGMP mediated, indicating alternative mechanisms of action, including nitrosylation of various target proteins. Thus, NO is a major controller of MC phenotype and function, inhibiting many allergy-associated activities and enhancing other non-allergy associated responses. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05020t1.jpg] [oc05020t2.jpg] |
| |||||||||

{kind=link}
{kind=link}