 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
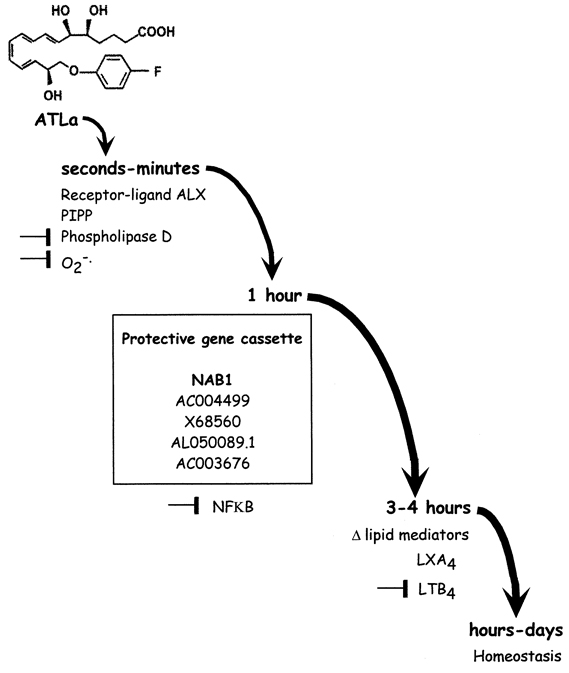
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol.100, Suppl. 1, March, 2005, pp. 55-57 Lipoxin agonists: turn right! to path of resolving neutrophil Pallavi R Devchand Center for Excellence in Vascular Biology, Cardiovascular Division, Department of Medicine, Harvard Medical School and Brigham and Women's Hospital, NRB 742 Boston, MA 02115, US. E-mail: pdevchand@rics.bwh.harvard.edu Received 8 November 2004 Code number: oc05027 An impressive array of cellular and molecular adaptive responses achieves homeostasis. The inflammatory reaction is an adaptive response triggered by an insult to culminate into the overt cardinal signs of inflammation, eventually leading to resolution and returning the organism back to its original centered state. This article focuses on some aspects of the lipoxin A4 signaling pathway during the resolution phase, to better understand molecular mechanisms by which a neutrophil directs an inflammatory reaction to switch off and resume homeostasis. Defining the resolution state of a neutrophil at the molecular level will aid in treatments of diseases that are associated with an exaggerated and uncontrolled inflammation. Key words: neutrophil - lipid mediators - resolution of inflammation Some of us would like to experience the landmark age of a hundred. More importantly, we do not just want to live longer; we want full, functional and active lives. So the challenge of that century, is to adapt to an ever-changing environment while maintaining identity and integrity. If we take that mission statement to the process of adaptive homeostasis during an acute inflammation, the cellular and molecular mechanisms that persistently bring the organism back to a centered state can be defined as resolution. The function of neutrophils (PMNs) is pivotal to the outcome of an acute inflammatory response. These cells respond to and generate many lipid and protein mediators that exert pro- and anti-inflammatory effects. Much attention has focused on the pro-inflammatory molecular aspects of these cells. However, little is known about the mechanisms associated with control and resolution of inflammation. LIPOXYGENASE PRODUCTS IN INFLAMMATION Major advances have been made in understanding signaling by arachidonic acid-derived lipid mediators (for review see Samuelsson et al. 1987). Eicosanoids are generated in small amounts (nano- to mico-molar quantities) and are rapidly inactivated. Even though these autocoids are short-lived, they are potent stimulators of bioactivity. A retrospective view of eicosanoid research emphasizes the importance of a multidisciplinary approach coupled to the use and development of stable synthetic mimetics. Three major lipoxygenase (LOs) have been cloned from human tissues (Funk 2001). These enzymes catalyse the production of lipid mediator precursors from arachidonic acid. For example, the 5-LO cascade leads to formation of leukotriene B4 (LTB4). Sequential oxygenation of arachidonic acid by lipoxygenases (eg. at C-15 and then C-5 position by 15-LO and 5-LO, respectively) results in the formation of lipoxygenase interaction products (lipoxins; Serhan et al. 1997). Lipoxins (LXs) are characterized structurally by the presence of four conjugated double bonds. Interestingly, the presence of aspirin during an inflammatory response, can result in the production of 15-epi-LXA4, also known as aspirin-triggered lipoxin (ATL). LESSONS IN ANTI-INFLAMMATORY MECHANISMS FROM LTB4 AND LXA4 SIGNALING PATHWAYS Studies on LTB4 and LXA4 signaling pathways have contributed key conceptual frameworks to our understanding of inflammation control. It is clear that resolution of an inflammatory reaction occurs at many levels of signal transduction and involves crosstalk between multiple pro-inflammatory and anti-inflammatory pathways. In PMNs, some key protective cellular mechanisms that exert counter-regulation of pro-inflammatory signals are attributed to: inhibition of signaling from cell surface receptors (Takano et al. 1997); clearance of lipid mediators (Devchand et al. 1996); inhibition of pro-inflammatory transcription factor activation pathways (Delerive et al. 1998); metabolic conversion of pro-inflammatory lipid mediators to anti-inflammatory mediators (Serhan 1997); and modulation of the chemokine-cytokine axis (Hachicha et al. 1999). More recent efforts have identified a nuclear co-repressor as an inducible switch associated with a resolution program, consistent with the notion that a "resolving neutrophil" has its own fingerprint of expressed genes (Qui et al. 2001). LX A4 ACTIONS LX A4 is generated mainly by transcellular biosynthesis during cell to cell interactions (Serhan 1997). The development of stable synthetic LXA4 and ATL mimetics has greatly facilitated studies on LXA4 functions and targets (Serhan 1997). Since these synthetic compounds are less susceptible to metabolic inactivation and appear to be functionally equivalent to LXA4, they have proven instrumental in the identification of novel mechanisms leading to the resolution of an inflammation. Based on in vivo and in vitro studies, we can now associate immu-nomodulatory and vasoactive protective roles with LXA4. For instance, in vivo ATLa inhibits PMN-trafficking in response to direct pro-inflammatory stimuli, as well as in second organ damage associated with ischemia-reperfusion injury (Qui et al. 2000). ATLa can also redefine the chemokine-cytokine axis in human PMNs, to downregulate the expression of pro-inflammatory peptides and upregulate the expression of anti-inflammatory peptides (Hachicha et al. 1999). For example, in human PMNs, ATLa decreases levels of TNFa-stimulated IL1b transcripts. Co-transfection assays in HEK 293 cells delineate this ATLa-stimulated reduction in cytokine production in part, as a receptor-mediated suppression of NFkB activity in the nucleus (Devchand et al. 2003). LIPOXIN A4 RECEPTORS OF THE GPCR CLASS Interactions of signaling molecules with cell surface receptors involves activation of downstream secondary signaling messengers including Ca2+, inositol phosphates, cAMP, G proteins and various kinases to eventually modulate gene expression (for review see Gether & Kobilka 1998). These secondary pathways have been studied extensively in context of pro-inflammatory stimuli. The signaling events associated with anti-inflammatory mediators are of great therapeutic interest. One emerging concept is the involvement of cell surface GPCRs in mediating the counter-regulation of pro-inflammatory signals (e.g. LTB4) by anti-inflammatory signals (e.g. LXA4), ultimately leading to inhibition or resolution of inflammation (for review see Serhan 1997). The cDNA for a LXA4 receptor (ALX) has been cloned from several vertebrates including mouse and man (Perez et al. 1992, Fiore et al. 1994, Takano et al. 1997). Sequence alignment indicates that the gene is well conserved with a 78% homology at the nucleic acid level and 73% identity at the amino acid level. The predicted protein structure is a seven transmembrane domain protein characteristic of the G protein-coupled receptor (GPCR) superfamily. Northern blot analyses indicate myeloid-selective expression, with striking abundance in human PMNs (Takano et al. 1997). ALX is a high affinity receptor for LXA4 (Kd of ~1.5 nM) that responds to LXA4 as a GPCR (Takano et al. 1997). When evaluated in vitro for activity, ALX also selectively binds and responds to synthetic LXA4 and ATL analogues (Takano et al. 1998). Recent experiments in mice transgenic for the human ALX (hALX) demonstrate that hALX is a cell surface receptor for the xenobiotic ATLa, as measured by inhibition of zymosan-stimulated neutrophil infiltration in a peritonitis model (Devchand et al. 2003). GAIN-OF-FUNCTION MOUSE MODEL FOR RESOLUTION In a proof-of-concept experiment, a transgenic (Tg) mouse model of selective expression of the human ALX in mature myeloid cells was used to demonstrate that ALX is a cell surface receptor that modulates dynamics of acute inflammation to trigger early resolution of inflammation (Devchand et al. 2003). Since ALX is a ligand activated receptor, exploiting the gain-of-function phenotype the hALX-Tg mouse offers a humanized pharmacogenetic system to elucidate dynamic regulation of endogenous mechanisms of resolution. Analyses of zymosan-induced exudates during progression of the acute inflammatory response revealed that hALX influenced the composition of soluble factors released in the inflammatory milieu, as measured by potency of the soluble fraction in an ex vivo bioassay that quantifies activation of the inflammation biomarker NFkB. Consistent with this, lipidomic analyses revealed decreased levels of LTB4 in Tg versus non-Tg wild-type littermates. Interestingly, the Tg mice revealed altered product profiles of the lipoxygenase signaling axes, including amplified production of LXA4. These in vivo data offer definitive proof that resolution of acute inflammation is an active program with genetic determinants that can be effectively modulated by signaling through the lipoxin receptor, ALX. TEMPORAL MAP OF THE ATLa SIGNALING PATHWAY IN THE NEOTRPHIL In the dynamic complexity of acute inflammation and host-defense, the outcome can be influenced to a large extent by two main classes of molecules: the signals generated and the receptors that receive and transmit them. In vivo, amplifying of the ALX receptor circuit can have a downstream protective impact on signal generation to reduce the number of PMN infiltrating into tissues and modulate the inflammatory set-point. Thus, it appears that ATLa, like all conventional signaling molecules, triggers a specific signaling circuit with temporal precision and amplitude (Figure) to influence the desired successful outcome for the host: adaptive homeostasis. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05027f1.jpg] |
| |||||||||

{kind=link}