 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol.100, Suppl. 1, March, 2005, pp. 113-120 Mechanisms of leukocyte lipid body formation and function in inflammation Patrícia T Bozza+, Christianne Bandeira-Melo Laboratório
de Imunofarmacologia, Departamento de Fisiologia e Farmacodinâmica,
Instituto Oswaldo Cruz-Fiocruz, Received 8 November
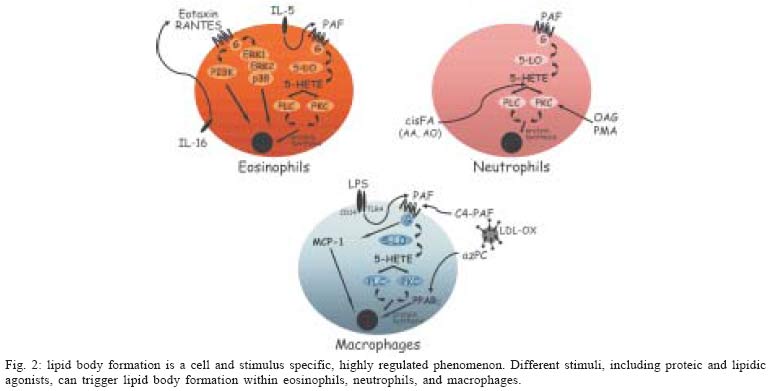
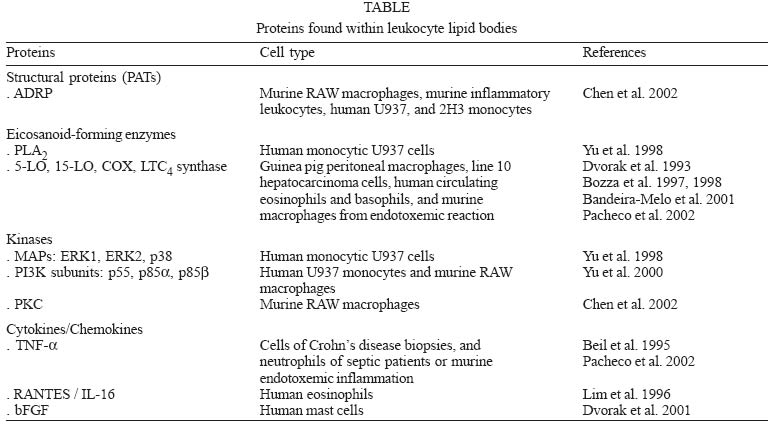
2004 Code number: oc05037 An area of increasingly interest for the understanding of cell signaling are the spatio-temporal aspects of the different enzymes involved in lipid mediator generation (eicosanoid-forming enzymes, phospholipases and their regulatory kinases and phosphatases) and pools of lipid precursors. The compartmentalization of signaling components within discrete and dynamic sites in the cell is critical for specificity and efficiency of enzymatic reactions of phosphorilation, enzyme activation and function. We hypothesized that lipid bodies - inducible non-membrane bound cytoplasmic lipid domains - function as specialized intracellular sites of compartmentalization of signaling with major roles in lipid mediator formation within leukocytes engaged in inflammatory process. Over the past years substantial progresses have been made demonstrating that all enzymes involved in eicosanoid synthesis localize at lipid bodies and lipid bodies are distinct sites for eicosanoid generation. Here we will review our current knowledge on the mechanisms of formation and functions of lipid bodies pertinent to inflammation. Key words: inflammation - lipid bodies - eicosanoids The cells of both innate and adaptive immune response express a great variety of receptors that transduce and integrate an enormous amount of information enabling the cells to mount effective immune responses. Over the past several years major advances have been made to elucidate the biochemical and molecular details of cell activation and depicting signaling cascades that leads to inflammatory mediator production. However the temporal and spatial organization of signaling and its impact on cell function is less clear. Recent evidence indicates that immune receptors and components of their signaling cascade are spatially organized within the cell and this spatial organization plays a key role in the regulation and amplification of cell signaling. Many studies have delineated roles for lipid domains including plasma membrane-associated domains - lipid rafts and caveolae and intracellular lipid-rich domains - named lipid bodies -, in cellular signaling. The functions of rafts and caveolae in cell signaling have been recently reviewed elsewhere (Anderson & Jacobson 2002, Dykstra et al. 2003, Pike 2003, van Deurs et al. 2003, Helms & Zurzolo 2004). Cytoplasmic lipid bodies are osmiophilic organelles present in virtually all mammalian cells, which are surrounded by a monolayer of phospholipids with a unique fatty acid composition, have a neutral lipid-rich core, and present variable protein composition (Murphy 2001, van Meer 2001, Tauchi-Sato et al. 2002). Members of the PAT family of proteins [perilipin, adipose-differentiation related protein (ADRP) and TIP 47], in particular ADRP, have been ubiquitously associated to lipid bodies in different cell types, including leukocytes, and although their function are not fully understood, they seem to act as structural proteins surrounding lipid bodies serving as nucleation center for lipid assembling and/or acting as an anchor for subcellular locations within the cell (Heid et al. 1998, Nakamura & Fujimoto 2003, Brasaemle et al. 2004). It is hypothesized that lipid body lipidic and proteic composition varies according to the cell type and stimulatory conditions, with impacts to their function in the cell. Here we review the evidence that leukocyte lipid body formation and compartmentalization of enzymes within lipid bodies is a highly regulated cellular event and that lipid bodies play a key role in the heightened capacity of leukocytes to generate eicosanoids in inflammatory conditions. MECHANISMS OF LEUKOCYTE LIPID BODY FORMATION Lipid bodies, although small in number, are normal constituent of leukocytes, including eosinophils, neutrophils, and monocyte/macrophages. Although lipid bodies, lipid-enriched organelles often overlooked if their defining lipid content is lost during cell staining, have long been noted to be prominent in many cell types associated with inflammation, the genesis and function of these organelles are still not completely defined. Indeed, most routinely used hematological staining protocols including May-Grünwald-Giemsa staining cause dissolution of lipid bodies (Fig. 1b, d), which precludes their recognition in stimulated leukocytes. However, with appropriate lipid fixation and staining, increased lipid body numbers have been observed in activated leukocytes as after LPS administration in vivo (Fig. 1c), compared with the control (Fig. 1a). Lipid bodies were shown to characteristically increase both in size and number in vivo in cells associated with human inflammatory diseases; including in blood eosinophils from patients with hypereosinophilic syndrome (HES) and in tissue eosinophils in biopsies from Crohn's disease (Solley et al. 1976, Beil et al. 1995, Bozza et al. 1998), in synovial leukocytes from patients with arthritis (Weinstein 1980, Schlesinger et al. 1982, Reginato et al. 1985, Bozza et al. 1996b) and lung and blood leukocytes from septic and/or ARDS patients (Triggiani et al. 1995, Pacheco et al. 2002). Genesis of new lipid bodies is not a manifestation of cellular injury or simply attributable to excess substrate fatty acids, on the contrary, it is a rapid, highly regulated phenomenon that is stimulus and cell specific (Fig. 2). For instance, lipid bodies can be elicited in vivo and in vitro after stimulation of cells with either lipid agonists such as, cis-unsaturated fatty acids, PAF, LPS or PKC activators (Weller et al. 1989, 1991 Bozza et al. 1996a, b, Pacheco et al. 2002) or protein stimuli such as chemokines and interleukines (Bozza et al. 1998, Bartemes et al. 1999, Bandeira-Melo et al. 2001a, 2002a). Neutrophil lipid bodies, identical to lipid bodies in leukocytes in vivo as evidenced by light and electron microscopy (EM), can also be rapidly (within 15-60 min) elicited in vitro after stimulation with cis-unsaturated fatty acids, PAF, or PKC activators (Weller et al. 1989, 1991, Bozza et al. 1996a, b). Although incubation of leukocytes with exogenous phospholipids and fatty acids provides a source of lipids for incorporation into newly formed lipid bodies, mechanisms other than simple availability of lipid precursors seem to be involved in lipid body formation. First, specific unsaturated fatty acids, including non-esterifiable analogs, in a stereochemically-restricted manner, can elicit de novo formation of lipid bodies in human leukocytes, whereas fully saturated fatty acids are not active (Weller et al. 1989, 1991, Bozza & Weller 2001). Moreover, PAF but not its precursor and metabolite, lyso-PAF, that shares the lipid structure of PAF but has no receptor agonistic activity (Prescott et al. 2000), have the capacity to induce lipid body formation (Bozza et al. 1996a, 1997, de Assis et al. 2003). In addition, it has been demonstrated that PAF-induced lipid body formation in leukocytes involves the regulated activation of different signaling pathways (Fig. 1). PAF stimulated lipid body is a receptor-dependent phenomenon, inhibitable by PAF receptor antagonist and pertussis toxin (Bozza et al. 1996a, 1997, 1998). PAF, acting via its G-protein-linked receptor induces lipid body formation via a downstream signaling involving, sequentially, 5-LO activation to generate 5-HETE that per se activates its G-protein-linked receptor, and protein kinase C activation (Bozza et al. 1996a). Interestingly other neutrophil agonists IL8, C5a, and LTB4 acting through G protein coupled receptors were not capable of inducing neutrophil lipid body formation (Bozza et al. 1996a). Of special relevance to allergic inflammation, key mediators of the allergic response in addition to PAF are capable of eliciting newly formed lipid bodies within human eosinophils. IL-5 (a stimuli proteic in nature) alone or combined to GM-CSF in the absence of exogenous lipids as well as immobilized IgG lead to significant increase in lipid body numbers (Bozza et al. 1998, Bartemes et al. 1999), suggesting that receptor mediated stimuli results in intracellular lipid remodeling and lipid body formation. Moreover, chemokines acting via CCR3 receptors, including RANTES, eotaxin, eotaxin-2, and eotaxin-3, can initiate intracellular signaling in eosinophils, but not neutrophils, that also culminate in de novo formation of lipid bodies (Bandeira-Melo et al. 2001a, b). Differently from the signaling pathways described by PAF, eotaxin and RANTES activating CCR3 receptor signals through phospho-inositide 3-kinase (PI3K) and the ERK1/2 and p38 MAP kinases (Bandeira-Melo et al. 2001a). Increased lipid body formation in neutrophils and macrophages could also be observed in experimentally induced inflammation triggered either by LPS or OxLDL (oxidized low density lipoprotein) but not by native LDL (Pacheco et al. 2002, Silva et al. 2002). LPS administration into mice induced a dose- and time-dependent increase in lipid body numbers. Moreover, LPS failed to form lipid bodies in C3H/HeJ (TLR4 mutated) mice or in cells treated with neutralizing anti-CD14, demonstrating a requisite role for LPS receptors in lipid body formation (Pacheco et al. 2002). Infection by T. cruzi also leads to increase in lipid body numbers in macrophages (Melo et al. 2003), whether this reaction would also involve pattern recognition receptors are now under investigation. PAF and PAF-like lipids are believed to play an important role in lipid body formation induced by LPS or by oxLDL in vivo since the pretreatment with three structurally unrelated PAF receptor antagonists completely inhibited this phenomenon (Pacheco et al. 2002, Silva et al. 2002). Although PAF may act at intracellular binding sites to induce cell activation (Bazan et al. 1994), results are suggestive that PAF is acting in a paracrine/autocrine way to induce lipid body formation, since the PAF-receptor antagonist used in those studies would act preferentially at membrane receptors and confirmed by findings that LPS or oxLDL induced lipid body formation were inhibitable by treatment with extracellular PAH-acetylhidrolase (Silva et al. 2002). It has been demonstrated that dietary fatty acids may have impacts on cellular lipid domains (Yaqoob 2003). Consumption of the long chain n-3 PUFA has been demonstrated to modify composition of lipid rafts and caveolae (Stulnig et al. 2001, Fan et al. 2003, 2004, Ma et al. 2004). We have recently demonstrated that fatty acid dietary intake may also modulate leukocyte lipid body formation. Indeed, dietary intake of extra virgin olive oil lead to inhibition of LPS-induced lipid body formation and decreased generation of MCP-1 and eico-sanoids (Leite et al. 2005). Interestingly, the pretreatment of leukocytes with protein synthesis inhibitors acting on transcription (actinomycin D) and translation (cycloheximide) significantly, although partially, inhibited lipid body formation induced by PAF and fatty acids in vitro but not by eotaxin and RANTES, thus indicating that according to the stimulated conditions induction of lipid bodies depends on new protein synthesis and its likely that specific and transcriptionaly regulated early response genes are activated during the process of lipid body formation (Bozza et al. 1996a, b, 1997, Bandeira-Melo et al. 2001a, Pacheco et al. 2002). A role for peroxisome proliferator-activated receptors (PPARs), members of the nuclear receptor supergene family that function in ligand-activated transcription, in leukocyte lipid body formation has recently been suggested. Indeed, we observed that treatment of mouse macrophages with BRL 49653, a preferential PPARg ligand, failed to induce lipid body formation. However, BRL 49653 treatment significantly potentiated lipid body formation induced by ox-LDL or PAF-like agonists, suggesting that PPAR have a role in regulating leukocyte lipid body formation (de Assis et al. 2003). However, we anticipate that differences in PPAR expression and role in lipid body formation may vary according to the cell type and stimuli studied and further studies are needed to characterize the roles of PPAR in lipid body formation and the involvement of PPAR targeted genes in this phenomenon. The formation of structurally distinct lipid bodies reflects mechanisms whereby proteins and lipids coalesce in a deliberate, highly regulated manner. Lipid body assembly can result from mobilization and reorganization of endogenously derived lipids as well as incorporating exogenous lipid. Thus, although lipid bodies are not conventional membrane-bound organelles, they appear to represent specialized intracellular domains whose induced formation is centrally related to activating mechanisms within the cells. Moreover, lipid body size and numbers are characteristically increased in leukocytes following activation in vitro as well as in in vivo inflammatory disorders, and can be used as a marker of leukocyte activation. LEUKOCYTE LIPID BODY FUNCTIONS IN INFLAMMATION Eicosanoid-forming enzyme compartmentalization - Several findings indicate a major role of lipid bodies in arachidonic acid metabolism during inflammation. First, lipid bodies are intracellular sites of arachidonate esterified within both neutral lipids (triacylglycerol and cholesterol) as well as in different phospholipid classes in cells including eosinophils, neutrophils and macrophages (Weller et al. 1989, 1991, Yu et al. 1998, Johnson et al. 1999). If lipid bodies are to have roles in eicosanoid mediator formation, then the arachidonic acid present in those lipid-rich structures must be released by phospholipases, and the free arachidonate must have access to eicosanoid-forming enzymes. Accordingly, the co-compartmentalization of cytosolic phospholipase A2 (cPLA2) and mitogen-activated protein (MAP) kinases, the upstream enzymes involved in AA liberation, within lipid bodies has been described (Yu et al. 1998). Immunocytochemistry from whole cell and immunoblotting of proteins from isolated lipid bodies (free of cytosol and other organelles) established that cPLA2 as well as the MAP kinases, ERK1, ERK2, p85, and p38, were localized at lipid bodies. In addition, isolated lipid body fractions contained cPLA2 enzymatic activity and released AA from sn-2-position of phosphatidylcholine (Yu et al. 1998). The two major enzymes, 5-LO and COX, involved in the enzymatic conversion of AA into eicosanoids were also shown to localize within lipid bodies. By using immunocytochemistry at the light level and ultrastructural postembedding immunogold, cyclooxygenase has been reported to localize at eosinophil lipid bodies, both in naturally formed lipid bodies in eosinophils from HES patients and in PAF-induced lipid bodies (Dvorak et al. 1992, 1993, Bozza et al. 1997, 1998). Previous studies on the intracellular localization of 5-LO have shown that 5-LO localization is cell type-specific and also varies according to the activation state of the cell. 5-LO was shown to localize within the nuclear environment (nuclear membrane and euchromatin) of alveolar macrophages and basophilic leukemia cells, whereas 5-LO was found to be predominantly cytosolic in human neutrophils and resting peritoneal macrophages (Peters-Golden & Brock 2000, Bandeira-Melo & Weller 2003). In addition to the nuclear environment, we have demonstrated the compartmentalization of the key enzyme for leukotriene production, 5-LO, within human eosinophil and basophil lipid bodies (Bozza et al. 1997, 1998, Bandeira-Melo et al. 2001a). In addition, im-muno-reactivity for 5-LO was also detected in lipid bodies from enucleated human eosinophils (Bozza et al. 1997). Human eosinophils upon stimulation preferentially produce LTC4 as their 5-LO product. LTC4-synthase is the terminal LT-forming enzyme and is present selectively in eosinophils, basophils, and mast cells (Bandeira-Melo et al. 2002a). Similarly to that observed for 5-LO, LTC4-synthase was shown to co-localize within eosinophil lipid bodies (Bozza et al. 1997). Sub-cellular localization of eicosanoid forming enzymes in leukocytes in vivo is starting to be unveiled. Li-pid bodies, in both ox-LDL- and LPS-induced models of inflammation, compartmentalize 5-LO and COX-2 enzymes within recruited leukocytes (Pacheco et al. 2002, de Assis et al. 2003). Similarly, eosinophils attracted to the site of allergic reaction showed 5-LO and COX-2 immuno-reactivity within newly formed lipid bodies. In addition, lipid bodies within leukocytes obtained from clinical inflammatory conditions including sepsis and arthritis were also demonstrated to be domains for intracellular compartmentalization of 5-LO and COX-2 (Pacheco et al. 2002, Roimicher, unpub. data). Together these findings support a role for lipid bodies to function as specific sites for eicosanoid formation. The compartmentalization of arachidonate substrate, cPLA2, and eicosanoid-forming enzymes provides in one locale an efficient mean to regulate arachidonate release and directly couple it with the enzymes to form eicosanoids (Table). Involvement of lipid bodies in enhanced generation of eicosanoids by inflammatory cells - Because leukocyte lipid bodies are sites of intracellular localization of eicosanoid-forming enzymes and also stores of the eicosanoid precursor arachidonic acid, it has been hypothesized that increases in lipid body numbers in leukocytes would result in enhanced capacity of eicosanoid production by leukocytes. Indeed, stimuli known to prime leukocytes to induce eicosanoid generation, including PKC activators, arachidonate and PAF, are also active in stimulating lipid body formation. Accordingly, others and we observed a significant correlation between lipid body formation and enhanced generation of both LO- and COX-derived eicosanoids in vitro (Bozza et al. 1996a, b, 1997, 1998, Bartmenes et al. 1999) as well as in vivo (Pacheco et al. 2002, Silva et al. 2002, de Assis et al. 2003, Melo et al. 2003). Analogously, agents that inhibited lipid body formation also resulted in inhibited priming for eicosanoid production. Indeed, lipid body formation induced by unsaturated fatty acids is inhibited by non-steroidal anti-inflammatory drugs (NSAIDs) including aspirin, indomethacin and, the non-cyclooxygenase inhibitor, sodium salicylate (Bozza et al. 1996b). Moreover, these NSAIDs block the priming response for increased prostanoid and leukotriene formation through a mechanism independent of their effects on cyclooxygenase but correlated with their capacity to inhibit lipid body formation (Bozza et al. 1996b, 2002). In addition, pretreatment of granulocytes with the protein synthesis inhibitors actinomycin D or cycloheximide inhibited not only PAF-induced lipid body formation, but also priming for LTC4 and PGE2 release by leukocytes, under conditions where they failed to inhibited calcium ionophore-induced LTC4 and PGE2 in cells not prestimulated with PAF (Bozza et al. 1996a, 1997). In order to evaluate if lipid bodies are sites involved in enhanced eicosanoid formation independently of the nuclear-pool of eicosanoid-forming enzymes and lipids, eosinophil enucleated cytoplasts were studied. As observed with intact eosinophils, PAF-induced a dose-dependent increase in the number of lipid bodies in eosinophil cytoplasts. Likewise, PAF-induced lipid body formation in nuclei-free cytoplast strongly correlated with increased LTC4 and PGE2 production following submaximal stimulation with A23187 (Bozza et al. 1997). That lipid bodies function as distinct extranuclear sites for eicosanoid formation was confirmed by the direct intracellular localization of newly formed eicosanoid. Direct assessment of intracellular sites for eicosanoid has been elusive, as those mediators are newly formed, non-storable and rapidly released upon cell stimulation. Recently, a new strategy to cross-link newly formed LTC4 at its sites of synthesis within in vitro stimulated eosinophils was described, enabling the immunofluorescent localization of newly formed LTC4 at its intracellular formation locale (Bandeira-Melo et al. 2001a, 2002b). Lipid bodies were the predominant sites of LTC4 synthesis in chemokine-stimulated human eosinophils. Therefore, CC chemokines elicit the formation of lipid body domains and promote LTC4 formation at these specific sites. Lipid bodies are also newly recognized sites of synthesis eico-sanoids that act as intracrine mediators regulating the release of a major cytokine, IL-4, stored within eosinophil granules (Bandeira-Melo & Weller 2005, in this issue). Collectively these findings indicate that lipid bodies are cytoplasmic, nuclei independent, sites for eicosanoid-forming enzyme localization, and eicosanoid production. Taken together with the evidences indicating the nucleus environment as important pools for eicosanoid metabolism (Serhan et al. 1996, Peters-Golden & Brock 2000, Bandeira-Melo & Weller 2003), raises intriguing possibilities that different sites for eicosanoid production might exist in a cell leading to the production of eicosanoids with intracrine function on regulation of transcription or regulating degranulation and for external release as para-crine inflammatory mediators. Putative lipid body functions based on protein compartmentalization - Recent studies focusing on the protein profile of lipid bodies in different cell types have revealed how broad lipid body functions can be. Among a growing list of proteins found within lipid bodies, it is known that they compartmentalize fatty acid metabolic enzymes, caveolin, kinases, small GTPases and cytokines, and therefore lipid bodies may regulate lipid metabolism, membrane trafficking, intracellular signaling and cell to cell communication. Although not yet characterized in leukocytes, lipid bodies from different cell types are sites of localization of lipid metabolic enzymes, including key enzymes involved in cholesterol metabolism (squalene epoxidase, 17-b-hydroxysteroid dehydrogenase, lanosterol synthase) and key enzymes of fatty acid synthesis (acetyl-coenzyme A carboxilase, NADH cytochrome b5 reductase), suggesting that both anabolic and catabolic steps in lipid metabolism may take place at lipid bodies (Brasaemle et al. 2004, Fujimoto et al. 2004, Liu et al. 2004, Umlauf et al. 2004). Proteins potentially associated with lipid trafficking including the cholesterol-binding protein caveolin and stomatin may be targeted to lipid bodies according to stimulatory conditions (Fujimoto et al. 2001, Ostermeyer et al. 2001, Pol et al. 2001, 2004, Umlauf et al. 2004). Moreover, the finding that cytosolic phospholipase A2 (cPLA2) and mitogen-activated proteins (MAP) kinases (also known as extracellular signal-regulated kinases [ERKs]), protein kinase C (PKC) and phosphatidylinositide 3-kinase (PI3K) - key enzymes implicated in intracellular lipid signaling of diverse cellular responses - are present at leukocyte lipid bodies suggests that regulatory signal transduction responses occur at lipid body domains (Yu et al. 1998, 2000, Chen et al. 2002) (Table). Of interest to the functions lipid bodies may have in inflammation, it has been demonstrated that cytokines, chemokines, and growth factors can be localized intracellularly in lipid bodies formed in activated leukocytes (Table). TNF-α was detected by an immunogold ultrastructural morphometric approach within cytoplasmic lipid bodies of several cell types, including fibroblasts, eosinophils, macrophages, mast cells, colonic epithelial cells and neutrophils in colonic Crohn's disease biopsies (Beil et al. 1995). TNF-α, were also shown to co-localize within leukocyte lipid bodies formed after LPS stimulation in vivo and within lipid bodies from peripheral blood neutrophils and monocytes from septic patients (Pacheco et al. 2002). Cytoplasmic lipid bodies of isolated human lung mast cells contain the basic fibroblast growth factor (bFGF), also found within mast cell secretory granules (Dvorak et al. 2001). Moreover, lipid bodies within human eosinophils are sites of localization of preformed RANTES and IL-16 (Lim et al. 1996), which are important eosinophil and lymphocyte activating agonists, and thus lipid bodies may have functions in allergic inflammation beside its role in LTC4 generation. Whether and how cytokines are released from lipid bodies, and whether cytokines have signaling functions within lipid bodies are questions that remain to be addressed. CONCLUDING REMARKS This review presents leukocyte lipid bodies as specialized, inducible intracellular domains that function as signaling platforms in inflammatory mediator production in the sense that the compartmentalization of substrate and key enzymes within intracellular lipid bodies have direct impact on the capacity of activated leukocytes to generate increased amounts of eicosanoids. Moreover, lipid bodies constitute sites of intracellular cytokine localization and may have functions beyond arachidonic acid metabolism. The emerging role of lipid bodies as inflammatory organelles raises lipid body status to critical regulators of different inflammatory diseases and key markers of leukocyte activation. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05037f2.jpg] [oc05037t1.jpg] [oc05037f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}