 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol.100, Suppl. 1, March, 2005, pp. 145-151 Regulation of stem cell factor expression in inflammation and asthma Carla A Da Silva, Nelly Frossard+ Faculté de
Pharmacie, Université Louis Pasteur, BP 24, 67401 Illkirch cedex,
France Received 8 November
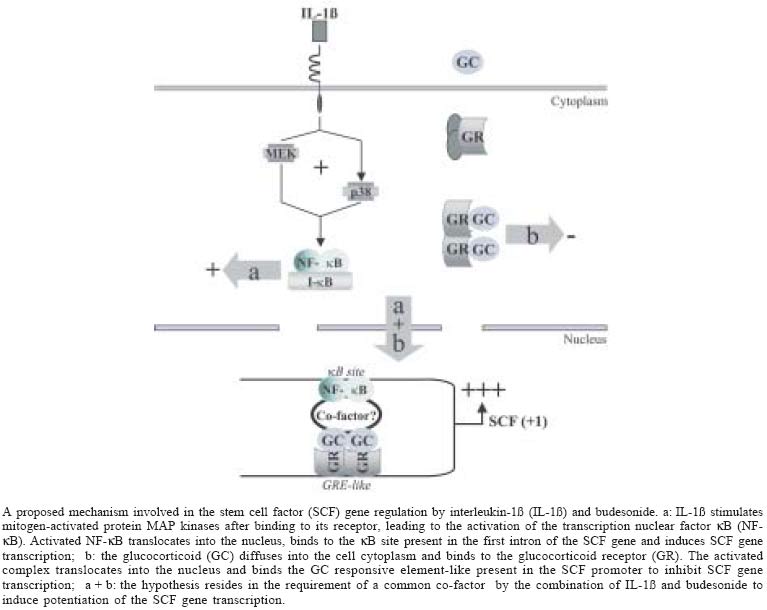
2004 Code number: oc05042 Stem cell factor (SCF) is a major mast cell growth factor, which could be involved in the local increase of mast cell number in the asthmatic airways. In vivo, SCF expression increases in asthmatic patients and this is reversed after treatment with glucocorticoids. In vitro in human lung fibroblasts in culture, IL-1ß, a pro-inflammatory cytokine, confirms this increased SCF mRNA and protein expression implying the MAP kinases p38 and ERK1/2 very early post-treatment, and glucocorticoids confirm this decrease. Surprisingly, glucocorticoids potentiate the IL-1ß-enhanced SCF expression at short term treatment, implying increased SCF mRNA stability and SCF gene transcription rate. This potentiation involves p38 and ERK1/2. Transfection experiments with the SCF promoter including intron1 also confirm this increase and decrease of SCF expression by IL-1ß and glucocorticoids, and the potentiation by glucocorticoids of the IL-1ß-induced SCF expression. Deletion of the GRE or kB sites abolishes this potentiation, and the effect of IL-1ß or glucocorticoids alone. DNA binding of GR and NF-kB are also demonstrated for these effects. In conclusion, this review concerns new mechanisms of regulation of SCF expression in inflammation that could lead to potential therapeutic strategy allowing to control mast cell number in the asthmatic airways. Key words: stem cell factor - inflammation - asthma The stem cell factor (SCF), also called Kit ligand, steel factor or mast cell growth factor (Huang et al. 1990, Martin et al. 1990, Zsebo et al. 1990), is the ligand of the product encoded by the proto-oncogene c-kit. SCF is expressed in two forms, soluble (sSCF) and membrane-bound (mSCF) after alternative splicing of the sixth exon, which encodes a proteolytic cleavage site (Anderson et al. 1991, Flanagan et al. 1991). SCF is involved in the early phases of hematopoiesis (de Vries et al. 1991, Brandt et al. 1992) (for review, see Galli et al. 1994, Broudy 1997). In particular, this growth factor also acts as an important growth factor for human and murine mast cells (Galli et al. 1994, 1995, Broudy 1997), including in vitro proliferation and differentiation of immature CD34+ progenitors into mast cells in the bone marrow (Kirshenbaum et al. 1992), and in peripheral blood (Rottem et al. 1994, Welker et al. 2000). SCF improves survival of mast cells by inhibiting their apoptosis (Mekori et al. 1993, Iemura et al. 1994). It induces their chemotaxis (Meininger et al. 1992, Nilsson et al. 1994), and plays a role in their adhesion to the extracellular matrix (Dastych & Metcalfe 1994, Kinashi & Springer 1994). In its membrane form, SCF acts as an adhesion molecule for them (Adachi et al. 1992, 1995, Kinashi & Springer 1994). SCF induces mast cell hyperplasia in vivo after subcutaneous injection in humans (Costa et al. 1996, Dvorak et al. 1998). In vitro it increases the antigen-induced degranulation of human pulmonary mast cells (Bischoff & Dahinden 1992, Okayama et al. 1994) and itself induces mast cell degranulation both in vitro (Columbo et al. 1992, Takaishi et al. 1994, Taylor et al. 1996a) and in vivo (Wershil et al. 1992, Costa et al. 1996). Mast cell number and activation are locally increased in chronic inflammatory diseases such as asthma (Lozewicz et al. 1988, Beasley et al. 1989, Djukanovic et al. 1990, Bradley et al. 1991, Ollerenshaw & Woolcock 1992, Saetta et al. 1992, Pesci et al. 1993, Laitinen et al. 1993, Bradding et al. 1994, Koshino et al. 1996). Since SCF is the principal growth factor for mast cells, it might be expressed in human bronchi and regulated in airway structural cells in pro- and anti-inflammatory conditions. That is, increased SCF expression in the bronchi of a patient with asthma would be associated with increased number and activation of mast cells. On the other hand, glucocorticoid anti-inflammatory treatment of asthma may diminish SCF expression and thereby reduce the number of mast cells. The present review relates studies of the expression and regulation of SCF expression in asthma conditions in vivo and in vitro. SCF EXPRESSION IN ASTHMA IN VIVO In vivo, SCF expression is regulated in the bronchi of asthmatic patients in both inflammatory and anti-inflammatory conditions, since this expression both increases and diminishes, respectively, together with the number of mast cells (Da Silva et al. 2004a). More particularly, two studies show that SCF expression in the bronchi is high in subjects with asthma, compared with healthy subjects (Da Silva et al. 2004a, Al-Muhsen et al. 2004). In addition, SCF expression is lower in glucocorticoid-treated subjects with asthma (Da Silva et al. 2004a). At the same time, the mast cell number is increased in the bronchi of asthmatic patients, often as degranulated, and decreased in the bronchi of asthmatic patients treated with glucocorticoids, where they eventually reached normal levels. These findings of the mast cell signature confirmed previous data from the literature in glucocorticoid-treated asthmatics (Bradding et al. 1994, Koshino et al. 1996, Djukanovic et al. 1997, Olivieri et al. 1997). Thus SCF, released by airway structural cells (Da Silva et al. 2002, 2003, 2004b, Kassel et al. 1998, 1999) could play some role in increasing the number and/or activation of mast cells in the human airways, and glucocorticoids may decrease mast cell number and activation at least in part through decreased SCF expression. Similar effects in human skin are suggested by work showing that glucocorticoids reduce the number of mast cells and the SCF expression (Finotto et al. 1997). SCF expression in the bronchi and the number of mast cells are regulated in pro- and anti-inflammatory conditions, but the mechanisms for this regulation are not yet known. This regulation of SCF expression has been studied in human airway cells in vitro to help understand how targeting SCF expression may help to treat asthma. SCF EXPRESSION IN PRO- AND ANTI-INFLAMMATORY CONDITIONS IN VITRO Effect of IL-1ß on SCF gene transcription - Interleukin (IL)-1ß, one of the major pro-inflammatory cytokine present within the asthmatic airways, increases the expression of SCF mRNA and SCF protein as a function of time (0 to 2.5 h), without modifying the relative expression of the two forms, sSCF and mSCF, at 30 min (Da Silva et al. 2004b). Studying the effect of inhibitors of the three mitogen-activated protein (MAP) kinase pathways showed that the MAP kinases p38 and extracellular signal regulated kinase (ERK)1/2 each accounted for 50% of the augmentation of SCF mRNA by IL-1ß at 30 min, while the Jun kinase pathway had no role in this effect (Da Silva et al. 2003). Chromatin immunoprecipitation experiments proved further that the transcription factor nuclear factor (NF)-kB binds at 30 min to a kB responsive element consensus sequence identified in the first intron of the SCF promoter gene (Da Silva et al. 2003). Deletion of this responsive element as well as pre-treatment with pyrrolidine dithiocarbamate (PDTC), a NF-kB inhibitor, shows that at 30 min NF-kB is the only transcription factor necessary for the early IL-1ß-induced increase in SCF expression (Da Silva et al. 2003, 2004b). Although other studies show that IL-1ß slightly increases SCF expression by structural cells at 24 h (Aye et al. 1992, Linenberger et al. 1995), less is known about the mechanisms involved in these effects. Interestingly, Da Silva et al. (2002) showed that SCF mRNA expression induced by IL-1ß peaked at 2.5 h and that it returned to normal levels within 24 h (unpublished observations). IL-1ß is known to stimulate the MAP kinase pathways within minutes (Lian et al. 1999), thereby stimulating such transcription factors as NF-kB (Janssen-Heininger et al. 1999, Liang & Gardner, 1999). The originality of the results by Da Silva et al. (2003) lies in the finding that a single transcription factor, located originally on an intron, transmits the effects of IL-1ß early (30 min) during this regulation of SCF expression (Da Silva et al. 2003). It is not yet clear if this remains true at later periods. Indeed, other responsive elements are present in the SCF gene promoter, in particular AP-1 or CREB (Taylor et al. 1996b), that might influence the effects of IL-1ß over a longer term, since reports show both of these transcription factors mediate effects of IL-1ß (Laporte et al. 2000, Song et al. 2003). SCF expression is thus increased in vitro by inflammatory conditions, as are other inflammatory mediators involved in asthma, such as eotaxin, the monocyte che-moattractant protein (MCP)-4 (Lamkhioued et al. 2000, Sato et al. 2001), and various interleukins. Accordingly, the inflammatory conditions created in vitro by IL-1ß treatment, which is responsible for the increase in SCF production, are consistent with the conditions that induce elevated SCF expression in vivo in the bronchi of subjects with asthma (Da Silva et al. 2004a, Al-Muhsen et al. 2004), associated with an elevated number and activation status of the mast cells in the airways (Da Silva et al. 2004a). Glucocorticoids and SCF gene transcription - Glucocorticoids, such as budesonide and dexamethasone, have been shown to diminish constitutive expression of SCF mRNA and SCF protein as a function of time (0 to 2.5 h), without modifying the relative expression of the two forms of SCF, sSCF and mSCF at 30 min (Da Silva et al. 2004b). This reduction was not modified by inhibitors of the three MAP kinase pathways, thereby confirming findings from the literature (Da Silva et al. 2003). These authors showed additionally by chromatin immunoprecipitation experiments that the glucocorticoid receptor (GR) complex binds to a GRE-like (glucocorticoid responsive element) sequence previously described in the SCF promoter (Taylor et al. 1996b, Da Silva et al. 2003, 2004b). Furthermore, transfection experiments with deletion of the GRE-like sequence and pre-treatment by RU486, the glucocorticoid receptor antagonist, showed that at short term (30 min) the effects of glucocorticoids on SCF expression are receptor-dependent. This inhibitory effect by glucocorticoids is consistent with the diminution in constitutive expression of SCF mRNA and protein at 2.5 h, previously observed (Kassel et al. 1998). The kinetics of the glucocorticoid-induced diminution in mRNA expression peaked at 2.5 h (Kassel et al. 1998, Da Silva et al. 2004b). Moreover, in a short time (30 min), the GR complex inhibited SCF expression by binding to GRE (Da Silva et al. 2003), which has been described as a transactivator element. The GRE sequence may overlap the sequence of another transcription factor, one involved in inhibiting basal SCF expression, but no responsive element corresponding to this hypothesis has so far been identified in the SCF promoter (Taylor et al. 1996b). Experiments with protein co-immunoprecipitation or chromatin immunoprecipitation might show whether GR is capable of interacting with other transcription factors, such as CREB or AP-1, which may, according to Taylor et al. (1996b), play a role in basal SCF expression. Glucocorticoids and IL-1ß-induced SCF gene transcription - The glucocorticoids, budesonide and dexamethasone, have been reported as surprisingly potentiating the IL-1ß-induced expression of SCF mRNA and protein at 30 min without modifying the relative expression of the two forms of SCF (30 min, Da Silva et al. 2004b). The effects of the three MAP kinase pathway inhibitors showed that p38 and ERK1/2 each accounted for 50% of this IL-1ß-induced potentiation of SCF mRNA at 30 min, while the Jun kinase pathway had no role then (Da Silva et al. 2003). This potentiation of SCF expression by the association of IL-1ß and glucocorticoids at 30 min is related to increased SCF gene transcription (Da Silva et al. 2004b), which in turn depends on the binding of NF-kB and of activated GR to their respective responsive elements (Da Silva et al. 2003). Moreover, the deletion of either of these responsive elements, or effect of the NF-kB inhibitor, PDTC, or of RU486 showed abolition of this potentiation, the induction of which therefore depends on an interaction between GR and NF-kB (Da Silva et al. 2003, 2004b). Very few studies in the literature report such synergy between the IL-1ß and glucocorticoid pathways or between NF-kB and GR. It has already been described for the cellular-inhibitors of apoptosis proteins, c-IAPs, for which NF-kB and dexamethasone have additive transac-tivator effects at 2 h, which are still greater at 24 h (Webster et al. 2002). No proposals so far attempt to explain the mechanisms involved in these effects. Altogether, these results suggest that a factor with co-activating properties may interact directly with NF-kB and GR (Da Silva et al. 2003). This factor must still be identified. For now, however, the direct interactions between NF-kB and GR have involved only a diminution in gene expression and not its stimulation or potentiation (Adcock 2001). For this reason, the hypothesis can be raised that factors may be needed to activate the bond between NF-kB and GR to potentiate SCF expression (Figure). The general transcription co-activator c-AMP binding protein (CBP) or the co-activator of the glucocorticoid receptor SRC (steroid receptor co-activator) might play a role in this effect. It is only after an hour of treatment that the expected diminution by budesonide of the IL-1ß-induced SCF expression is observed. Glucocorticoids are well known to inhibit the expression of mediators involved in inflammation, in particular, NF-kB. Many mechanisms for this inhibition of NF-kB by glucocorticoids have been proposed (for review, see Cato & Wade 1996). After the accumulation of GR in the cell, a protein-protein interaction may take place between NF-kB and GR (McKay & Cidlowski 1998) and prevent either the binding of NF-kB to its responsive element, or the transactivator effect of NF-kB associated with its responsive element (Nissen & Ya-mamoto 2000). Another possibility is the inhibition of the MAP kinase pathways by glucocorticoids, as suggested by the work of Kassel et al. (2001), who found that glucocorticoids induce the expression and activation of the MAP kinase phosphatase (MKP)-1. Additional experiments should confirm whether glucocorticoids inhibit the transactivator effect of NF-kB on SCF expression after 1 h, or if they inhibit the binding of other transcription factors, such as AP-1 (Diamond et al. 1990), to their responsive elements and the induction of their transac-tivator effects. In any case, the diminution by glucocorticoids of SCF expression in inflammatory conditions is consistent with the diminution of SCF expression reported in vivo, in subjects with asthma treated with glucocorticoids (Da Silva et al. 2004a). This is also true for other inflammatory mediators involved in asthma, such as MCP-4 (Lamkhioued et al. 2000). This diminution of SCF expression by glucocorticoids may contribute to the reduction in the number and activation status of mast cells in the airways of subjects with asthma after glucocorticoid treatment. SCF mRNA stability - Glucocorticoids and IL-1ß increase the stability of SCF mRNA very early (30 min), while neither IL-1ß nor glucocorticoids alone have any effect on this stability at such short term (Da Silva et al. 2004b). This stability is decreased by the association between IL-1ß and the glucocorticoids at 2.5 h, a time point at which IL-1ß alone increases SCF mRNA stability and where glucocorticoids alone diminish it (Kassel et al. 1998, Da Silva et al. 2002). Finally, the combination of glucocorticoids and IL-1ß also diminish the stability of SCF mRNA at 10 h, while neither IL-1ß nor glucocorticoids alone have any effect on this stability (Da Silva et al. 2002). The effect of IL-1ß alone on the increased stability of mRNAs has been described for cyclo-oxygenase (COX)-2 mRNA (Ridley et al. 1998). Glucocorticoids are also known to destabilize iNOS mRNA, whereas an increased stability of iNOS mRNA is reported at 15 h to be an effect of IL-1ß combined with dexamethasone (Kunz et al. 1996). Nonetheless, the signaling pathways leading to either stabilization or destabilization are not known. The presence of key sequences, the AURE (AU-rich responsive element) sequences in the 3' region of mRNA, generally determines whether IL-1ß and glucocorticoids stabilize or destabilize mRNA. SCF mRNA does not have an AURE sequence (Martin et al. 1990), however. This suggests that the mechanisms by which IL-1ß and glucocorticoids stabilize or destabilize SCF mRNA are still unknown. These mechanisms may involve regulation of the poly (A+) binding proteins to the mRNA (PABP) or of the polyadenylation process for mRNA, which would stabilize it (Stoeckle 1992, Ross 1995). Relative SCF expression - At short term treatment (30 min), the relative expression of the two forms of SCF, sSCF and mSCF, is not modified by IL-1ß, glucocorticoids or their combination. It is known that the relative expression of sSCF and mSCF varies in vitro according to cell type (Kassel et al. 1999) and that both forms can be involved in the augmentation of the number and/or activation of mast cells. Membrane-bound SCF, for example, may play a more important role than sSCF in the in vivo development of mast cells, as suggested by work in mutant mice homozygous for the Steel Dickie (Sld) allele: they produce biologically active sSCF but not mSCF and suffer from a profound mast cell deficiency (Brannan et al. 1991). mSCF also stimulates the development of hematopoietic cells still more than sSCF (Toksoz et al. 1992). A possible explanation is that the mSCF stimulation of the Kit receptor delays internalization of the SCF-Kit complex and accordingly delays the end of the stimulation (Miyazawa et al. 1991, Toksoz et al. 1992). Moreover, mSCF acts as an adhesion molecule for mast cells (Kinashi et al. 1994, Adachi et al. 1995). The relative expression of the two forms of SCF should be verified in patients with asthma who are and are not treated with glucocorticoids and compared with results in healthy subjects to see if expression of these two forms varies in vivo in humans during the disease. CONCLUSION In conclusion, SCF expression increases in pro-inflammatory conditions and diminishes in anti-inflammatory conditions in vitro. These findings confirm results obtained in vivo in patients with asthma. Glucocorticoids are also shown to have unexpected effects in the regulation of SCF gene at very short treatment times (30 min), since SCF expression is potentiated by glucocorticoids in pro-inflammatory conditions. The mechanisms involved in this effect are only partially elucidated. The hypothesis of the existence of a co-activator linking NF-kB and GR in the potentiation of SCF expression still needs to be confirmed. Once confirmed and identified, this co-activator might become a treatment target for inhibiting the harmful effect of SCF in disease. The physiological role of SCF in the airways and its role in asthma also require further elucidation. Altogether, SCF may be proposed as an interesting target for asthma treatment through its effect on the regulation of the number and activation status of mast cells, those inflammatory cells for which it is one of the most important growth factors. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz
The following images related to this document are available:Photo images[oc05042f1.jpg] |
| |||||||||

{kind=link}