 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol.100, Suppl. 1, March, 2005, pp. 177-180 Role of protease-activated receptor-2 in inflammation, and its possible implications as a putative mediator of periodontitis M Holzhausen/*, LC Spolidorio*, N Vergnolle+/++ Department of Pharmacology and Therapeutics, Faculty of Medicine, University of Calgary, Calgary, 3330 Hospital Drive, NW Calgary, T2N 4N1, Alberta, Canada *Departamento de Periodontologia e Patologia Oral, Escola Dental de Araraquara, Universidade Estadual de São Paulo, Araraquara, SP, Brasil Received 8 November
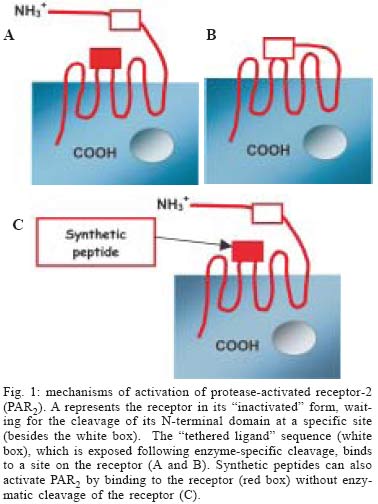
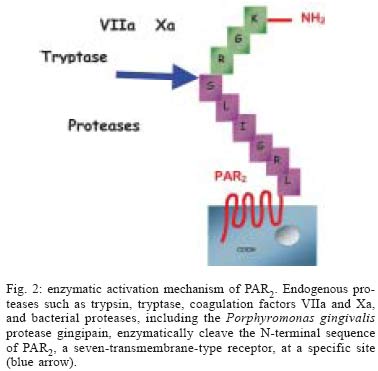
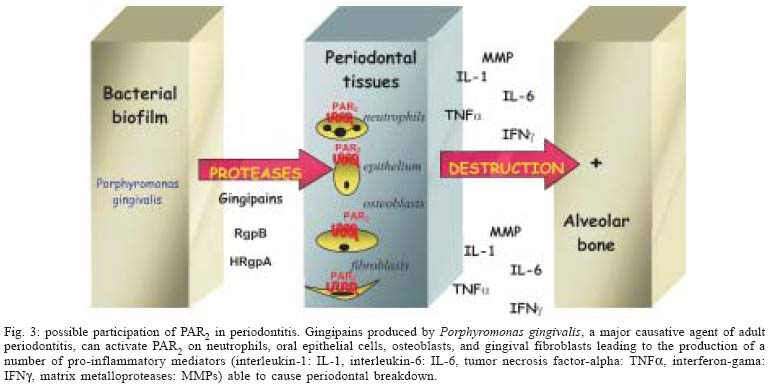
2004 Code number: oc05047 Proteinase-activated receptor-2 (PAR2) belongs to a novel subfamily of G-protein-coupled receptors with seven-transmembrane domains. This receptor is widely distributed throughout the body and seems to be importantly involved in inflammatory processes. PAR2 can be activated by serine proteases such as trypsin, mast cell tryptase, and bacterial proteases, such as gingipain produced by Porphyromonas gingivalis. This review describes the current stage of knowledge of the possible mechanisms that link PAR2 activation with periodontal disease, and proposes future therapeutic strategies to modulate the host response in the treatment of periodontitis. Key words: protease-activated receptor-2 - alveolar bone loss - inflammation - host response - Porphyromonas gingivalis - periodontitis Proteinase-activated receptors (PARs) belong to a recently described family of G-protein-coupled, seven-transmembrane-domain receptors. Activation of PARs occurs through proteolytic cleavage of their N-terminal domain by proteinases, resulting in the generation of a new N-terminal "tethered ligand", which can autoactivate the receptor function (see Figs 1A, B) (Ossovskaya & Bunnett 2004). Four members of the PAR family have been cloned. PAR1, PAR3, and PAR4 can be activated by thrombin, and PAR2 can be activated by trypsin, mast cell tryptase, neutrophil proteinase 3, tissue factor/factor VIIa/factor Xa, membrane-tethered serine proteinase-1, or proteases from Porphyromonas gingivalis (Fig. 2) (Vergnolle et al. 2001, Lourbakos et al. 2001). Selective synthetic peptides, corresponding to the tethered ligand sequences, are able to activate selectively the receptors through direct binding to the body of the receptor (Fig. 1C), without the need of proteolysis (Cocks & Moffatt 2000). With the exception of PAR3, all the other receptors have their selective agonist peptides. PAR1, PAR2, and PAR4 can be non-enzimatically and selectively activated by TFLLR-NH2, SLIGRL-NH2, and GYPGQV-NH2, respectively (Ossovskaya & Bunnett 2004). In spite of showing similar structures and common mechanisms of activation, the PARs have different tissue localization and function. PAR1 can be found in human platelets, endothelium, epithelium, fibroblasts, myocytes, neurons, and astrocytes, and it seems to play a role in the vascular matrix deposition after injury. PAR3 and PAR4 are found in platelets, endothelium, myocytes, and astrocytes, and they are thought to be involved in the thrombus formation and pulmonary embolism (Ossovskaya & Bunnett 2004). PAR2 is found throughout the body, especially in the epithelium, endothelium, fibroblasts, osteoblasts, neutrophils, myocytes, neurons, and astrocytes (Abraham et al. 2000, Uehara et al. 2003, Ossovskaya & Bunnett 2004). PAR2 seems to play critical pathophysiological roles, as it is involved in leukocyte migration, inflammation of joints, skin, and kidney and allergic inflammation of airways (Ossovskaya & Bunnett 2004). PAR2 AND INFLAMMATION The involvement of PAR2 in inflammation is supported by several studies. Early studies reported that PAR2 expression was up-regulated by inflammatory mediators such as tumour necrosis factor a, interleukin 1a and lipopolysaccharide (Nystedt et al. 1996). Furthermore, deletion of PAR2 also diminishes inflammation in the airway and joints (Schmidlin et al. 2002, Ferrell et al. 2003), and delays the onset of inflammation (Lindner et al. 2000). Moreover, a number of studies have demonstrated that activation of PAR2 can lead to blood vessel relaxation, hypotension, increased vascular permeability, granulocyte infiltration, leukocyte adhesion and margination, and pain (Cocks & Moffatt 2000, Vergnolle et al. 2001, Coughlin & Camerer 2003), all effects that encounter for the cardinal signs of inflammation. PAR2 activation also leads to the release of prostanoids and cytokines including interleukin IL-6 and IL-8 in epithelial or non-epithelial cells (Lourbakos et al. 2001, Uehara et al. 2003). In the gastrointestinal tract, PAR2 has been localized in many different cell types: in enterocytes, in endothelial cells of the lamina propria and the submucosa, in fibroblasts, in myenteric neurons, in immune and inflammatory cells (lymphocytes, neutrophils, mast cells) (Bohm et al. 1996, Nystedt et al. 1996). Recently, we have shown that in the colon, PAR2 agonists (PAR2-activating peptide, trypsin, tryptase) lead to an inflammatory reaction characterized by edema, granulocyte infiltration, increased intestinal permeability and pro-inflammatory cytokines (interleukin-1, TNF-α) release (Cenac et al. 2002). Recent studies also indicate an important role of PAR2 in inflammatory pain. The receptor identified on sensory afferent nerves has been associated with long-lasting thermal and mechanical hyperalgesia in the soma as well as in visceral organs (Vergnolle et al. 2001, Coelho et al. 2002). Taken together, these studies suggest a pro-inflammatory role for PAR2 in vivo, as it may mediate responses to tissue injury. These findings suggest that PAR2 plays a crucial role in the regulation of inflammation. ROLE OF PAR2 IN PERIODONTITIS A possible participation of PAR2 in chronic oral inflammation such as periodontitis was indirectly suggested by several studies. First, gingipain, a bacterial proteinase produced by P. gingivalis, a major causative agent of adult periodontitis, was reported to activate PAR2 (Lourbakos et al. 2001). In addition, PAR2 expression was found in osteoblasts, oral epithelial cells, and human gingival fibroblasts (Abraham et al. 2000, Lourbakos et al. 2001, Uehara et al. 2003). Lourbakos et al. (2001) showed that in an oral epithelial cell line, PAR2 activation by purified gingipain induced the secretion of the pro-inflammatory cytokine interleukin-6 (IL-6), which is a potent stimulator of osteoclast differentiation and bone resorption. Uehara et al. (2003) demonstrated that a synthetic PAR2 agonist peptide activates human gingival fibroblasts to produce IL-8 and to selectively stimulate MMP activity from these cells. This particular study suggests that PAR2 activation could account for collagen destruction associated with periodontitis lesions. Most recently, a study by Chung et al. (2004), showed that PAR2 is involved in the up-regulation of human beta-defensin in human gingival epithelial cells, stimulated by the peptide agonist of PAR2, and P. gingivalis proteases. Thus, this study points to a possible role for PAR2 in the gingival tissues, where its activation could act as an emergency mechanism, that would constitute a first alarm in mucosal tissues, alerting for the invasion of bacterial pathogens, and organizing a primary inflammatory response. Taken together, these studies suggest a role for PAR2 activation in inducing inflammation and bone resorption during periodontitis. However, another study by Smith et al. (2004) suggests that PAR2 activation could inhibit bone resorption. In that study, the authors showed that the selective PAR2-activating peptide SLIGRL-NH2 inhibited osteoclast differentiation, thereby acting as a potential inhibitor of bone destruction. This result, which contradicts the suggested role for PAR2 activation in bone loss, reflects the difficulties of using in vitro approaches to evaluate the role of the different mediators that are involved in periodontal diseases. The experiments from our group (data not published) provided the first evidences for in vivo evaluation of the role of PAR2 activation in periodontitis. We showed that local application of a selective PAR2 agonist (SLIGRL) in oral cavity of rats, causes gingival granulocyte infiltration, and periodontitis through a mechanism involving prostaglandin release and matrix metalloproteinase activation. In addition, seven days after PAR2-agonist treatment, a peak of granulocyte infiltration [measured by an increased myeloperoxydase (MPO) activity] was observed. As polymorphonuclear neutrophils represent the main source for MPO in acute inflammation, and because they constitute the frontline of the acute host inflammatory response, promoting the release of a number of inflammatory mediators that are able to stimulate osteoclasts (Dennison & Van Dyke 1997), it can be proposed that recruited neutrophils might be responsible, at least in part, for the initiation of periodontitis. Therefore, our study also suggests that PAR2 agonist-induced bone loss is due, at least in part, to the induction of an acute inflammatory response. In agreement with previous in vitro studies, which supported a destructive role for PAR2 (Uehara et al. 2000, Lourbakos et al. 2001), our in vivo approach definitively demonstrated a pro-inflammatory and bone destruction role for PAR2 activation in periodontal tissues. Proteinases, through the activation of PAR2, should then be added to the number of mediators implicated in periodontal diseases (Fig. 3). Interestingly, gingipains-R (RgpB and HRgpA) activate also the PARs, PAR1 and PAR4, which are expressed on the surface of platelets and are responsible for platelet aggregation (Lourbakos et al. 2001b). This mechanism may constitute the biological plausibility of the association between periodontitis and cardiovascular disease. However, no study has yet linked the role of PAR1 or PAR4 to periodontal diseases. CONCLUSIONS The pro-inflammatory role of PAR2 in inflammation is adequately and clearly demonstrated by several studies, which showed that PAR2 activation leads to widespread pro-inflammatory effects, including the release of pro-inflammatory cytokines, and regulation of a number of inflammatory diseases. The association of PAR2 with the pathogenesis of periodontitis is supported by some concepts: (i) PAR2 can be activated by gingipain, a bacterial protease produced by the major periodontopathogen, P. gingivalis; (ii) PAR2 is expressed by cells that are actively involved in periodontal pathologies, such as oral epithelial cells, fibroblasts, and osteoblasts, and PAR2 activation in those cells leads to the production of mediators of bone resorption; (iii) PAR2 activation by a selective peptide agonist leads to gingival granulocyte infiltration, and alveolar bone loss in rats, through a mechanism involving prostaglandin release and matrix metalloproteinase activation. These findings indicate that PAR2 might represent a potential target for the design of drug therapies focused on the modulation of periodontal inflammation. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05047f2.jpg] [oc05047f3.jpg] [oc05047f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}