 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
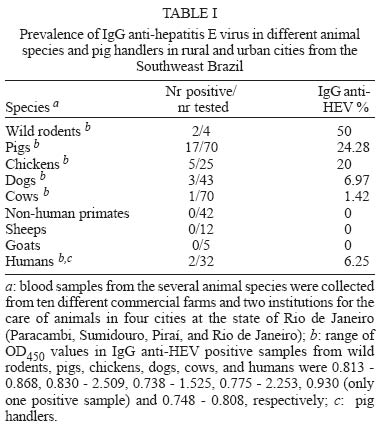
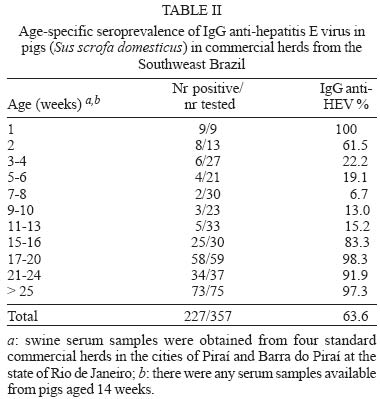
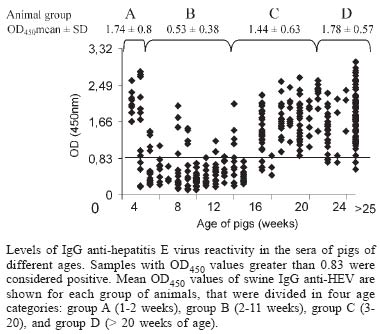
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 100, No. 2, April, 2005, pp. 117-122 Serological evidence of hepatitis E virus infection in different animal species from the Southeast of Brazil Cláudia L Vitral/*/+, Marcelo A Pinto, Lia L Lewis-Ximenez, Yuri E Khudyakov**, Débora R dos Santos, Ana Maria C Gaspar Departamento de Virologia, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21040-900 Rio de Janeiro, RJ, Brasil *Departamento de Microbiologia e Parasitologia, Instituto Biomédico, UFF, Niterói, RJ, Brasil **Division of Viral Hepatitis, WHO Collaborating Center for Research and Reference in Viral Hepatitis, Centers for Disease Control and Prevention, Atlanta, GA, US Financial support: Faperj, CNPq +Corresponding author. E-mail: clvitral@ioc.fiocruz.br Received 28 October 2004 Serological evidence of hepatitis E virus infection (HEV) has been observed in both humans and different animal species living in non-endemic areas, suggesting that animals could be important reservoir for virus transmission to man. Antibodies to HEV have been detected in some Brazilian population groups. Nevertheless, sporadic cases of acute HEV infection have never been reported. We collected 271 serum samples from several domestic animals and also from pig handlers from Southeast of Brazil in order to investigate the seroprevalence of HEV infection. Anti-HEV IgG was detected in cows (1.42%), dogs (6.97%), chickens (20%), swines (24.3%), and rodents (50%), as well as in pig handlers (6.3%). The recognition of swine HEV infections in pigs in many countries of the world led us to investigate a larger sample of pigs (n = 357) from the same Brazilian region with ages ranging from 1 to > 25 weeks. IgG anti-HEV was detected in 100% of 7-day old pigs. Following a gradual decline between weeks 2 and 8 (probably due to loss of maternal IgG), the prevalence then steady increased until it reached 97.3% of animals older than 25 weeks. Besides the detection of anti-HEV antibodies in different animal species, the results showed that swine HEV infection seems to be almost universal within this Brazilian pig population. This is the first report that shows evidences of HEV circulation in Brazilian animal species and pig handlers. Key words: anti-hepatitis E virus - animal reservoir - swine - Brazil Several aspects regarding hepatitis E virus (HEV) has led to a hypothesis that an animal reservoir for this virus exists, and that hepatitis E could be a zoonosis. First of all, when serological assays became available, a surprisingly high anti-HEV prevalence was found in blood donors and healthy individuals from non-endemic areas (Thomas et al. 1997, Mast et al. 1997, Meng 2000a,b). One of the possible reasons for this high seroprevalence could be the existence of animal reservoirs for HEV. This hypothesis was first experimentally examined by Balayan et al. (1990), who reported on the infection of domestic swine with a Central Asia strain of human HEV. Clayson et al. (1995) subsequently detected HEV RNA and anti-HEV IgG in pigs in Nepal, but the virus was not characterized. A unique swine HEV was first isolated in 1997 by Meng and collaborators. The swine HEV was shown to be genetically closely related to the human HEV. Afterwards, numerous strains of swine HEV have been genetically identified from pigs in both endemic and nonendemic regions of the world, showing the ubiquitous nature of swine HEV infection in pigs (Meng et al. 1997, 1999, Hsieh et al. 1999, Wu et al. 2000, Arankalle et al. 2001, Garkavenko et al. 2001, Okamoto et al. 2001, Huang et al. 2002). Moreover, recent seroepidemiological studies showed that swine veterinarians and other pig handlers are at higher risk of HEV infection compared to normal blood donors (Meng et al. 1999, 2002, Hsieh et al. 1999). Interspecies transmission of swine HEV has been experimentally demonstrated: swine HEV infected nonhuman primates, and a US strain of human HEV infected pigs (Meng et al. 1998a,b, Halbur et al. 2001). In addition to pigs, serological evidence of HEV infection was also documented in other animal species, such as rats, chickens, dogs, cows, sheep, goats, cattle and rhesus monkeys (Tien et al. 1997, Favorov et al. 1998, Kabrane-Lazizi et al. 1999, Arankalle et al. 2001). Finally, novel strains of human HEV recovered from acute hepatitis E cases have been recently reported and showed to be genetically more closely related to strains to swine HEV from respective countries than to other strains of human HEV (Hsieh et al. 1999, Wu et al. 2000, Takahashi et al. 2003). These observations suggest that these novel strains of human HEV may be of swine origin, and that swine HEV may infect humans. At the beginning of this study, no serologically confirmed acute cases of HEV infection have been documented in Brazil. However, anti-HEV antibodies were detected in a significant proportion of healthy individuals in the country (Pang et al. 1995, Foccacia et al. 1995, Parana et al. 1997). Our data show seroprevalence rates varying from 1 to 11.8% among blood donors, pregnant women, intravenous drug users and citizens from urban and rural areas (Souto et al. 1997, Oliveira et al. 1999, Trinta et al. 2001, Santos et al. 2002). In order to investigate the existence of animal reservoirs for HEV in Brazil, different animal species as well as pig handlers were tested for IgG anti-HEV. A larger collection of serum specimens from pigs of different age groups was also evaluated for the anti-HEV status. Results demonstrated that most animal species had anti-HEV antibodies. The HEV infection was found almost universal in swine by the age of 15 weeks. MATERIALS AND METHODS Collection of serum samples - A total of 271 blood samples were initially collected from several animal species from ten different commercial farms and two institutions for the care of animals (Center for the Breeding of Wild Animals and Jorge Vaistman Institute of Veterinary Medicine) in four cities at the state of Rio de Janeiro (Rio de Janeiro, Paracambi, Piraí, Sumidouro). Rio de Janeiro and Paracambi cities are located at the metropolitan region of the state, while Sumidouro and Piraí cities are situated at the mountain and medium Paraíba river regions, respectively. The following species were included: captive New World monkeys (Callithrix jacchus, C. kuhli, C. geoffroy, C. penicilata, C. argenta, Aotus sp.), dogs, cows, sheeps, goats, pigs, chickens, and small rodent (Nectomus sp.) as described in Table I. Additionally, serum specimens were collected from 32 pig handlers. A large collection of serum specimens (Sus scrofa domesticus) (n = 357) was obtained from pigs with ages ranging from 1 to > 25 weeks in four standard commercial herds in the cities of Barra do Piraí and Piraí, both localized at the medium region of the Paraíba river at the state of Rio de Janeiro (Table II). All serum samples used in this study were stored at -20°C prior to analysis. The protocols for specimen collection and use were submitted and approved by the Human Research Committee and the Animal Care Research Committee from Oswaldo Cruz Foundation, Health Ministery, RJ, Brazil. Detection of IgG anti-HEV - Serum samples were tested for IgG anti-HEV by EIA according to the method described previously and standardized by Obriadina et al. (2002) that uses two HEV recombinant proteins, a mosaic protein (MP-II) and a protein containing region 452-617 aa of the ORF2 of the HEV Burma strain as coating antigens. When applied to detection of HEV antibodies in animal sera, this EIA protocol was modified by using species-specific anti-IgG horseseradish peroxidase enzyme conjugates (anti-IgG-HRP, Sigma Chemicals St. Louis, MO) for serum samples representing respective species. Briefly, microtitre wells (Nalgen Nunc International, Denmark) coated with 100 µl of the recombinant HEV proteins (pB166 protein 1.2-1.4 µg/ml and MPII protein 1.1-1.2 µg/ml) dissolved in the 0.5 M carbonate buffer (CBB), pH 9.6, were incubated for 1 h at 37oC with 200 µl of 1% nonfat milk in CBB. Specimens were diluted 1/20 in PBS with 0.05% Tween 20 containing 5% normal goat serum (NGS), and 1% nonfat milk and incubated at 37oC for 30 min. After extensive washing, a volume of 100 µl of horseradish peroxidase labeled anti-animal species IgG or anti-human IgG (Sigma) appropriately diluted in PBST with 5% NGS and 1% nonfat milk was added to each well. The wells were incubated for 30 min at 37oC and washed 6 times, followed by the addition of 100 µl of substrate solution (tetrametylben-zidine and H2O2). After incubation for 15 min at room temperature in the dark, the enzyme reaction was stopped with 50 µl of 1 N H2SO4. The wells were read in an EIA reader set at 450 nm. The EIA cutoff for human serum samples was statistically established as the mean of the optical density (OD) value at 450 nm of known negative samples + 4SD of the mean. Due to the absence of known IgG anti-HEV negative serum samples for the different animal species evaluated, the cutoff values to define an initially reactive specimen were derived from the frequency distributions of optical density (OD) values obtained from the lowest reactivity IgG anti-HEV serum samples available from each of these animal species. The mean OD value at 450 nm of these specimens + 5SD was used as the cutoff value. In the case of swine IgG anti-HEV assay, 80 low reactivity IgG anti-HEV pig serum samples were evaluated. The OD values ranged from 0.038 to 0.493 and the value of 0.830, which was calculated as 5SD above the mean value (0.329), was used as the cutoff value. The specificity of EIA reactivity was evaluated by blocking assays. A sample was regarded as positive for IgG anti-HEV if a decreased to less than 30% of the original A450 value was observed after absorption with the same recombinant proteins used as antigen probe. Statistical analysis - In order to compare the frequency and the mean of OD450 values of swine IgG anti-HEV, animals were divided in four age categories: group A (1-2 weeks of age, n = 22), group B (3-11 weeks of age, n = 110), group C (12-20 weeks of age, n = 113) and group D (pigs older that 20 weeks of age, n = 112). Age categories were established after the calculation of the mean number of animals, with the exception of group A, that was composed only by animals with passive transferred IgG anti-HEV. Chi-square and unpaired F (Kruskal-Wallis) tests were applied using STATA (Intercooled Stata 6.0, Texas, US). A p value < 0.05 was considered statistically significant. RESULTS Prevalence of IgG anti-HEV in different animal species and pig handlers - IgG anti-HEV was detected in cows, dogs, chickens, pigs, and wild rodents with prevalence rates of 1.4, 7.0, 20.0, 24.3, and 50.0, respectively (Table I). Moreover, 2 out of 32 pig handlers (6.3%) were shown to be IgG anti-HEV positive. None of the nonhuman primates, sheep and goats tested positive for anti-HEV antibodies in their sera (Table I). Age-specific prevalence study of IgG anti-HEV in swine - IgG anti-HEV was detected in 100% of 7-days pigs (Table II). Following a gradually decline between weeks 2 and 8 (presumably loss of maternal IgG), a steady increased of the prevalence of IgG anti-HEV was initially observed from 13% of samples from pigs aged 9-10 weeks to 15.2% of animals aged 11-13 weeks. Then, the prevalence increased sharply to 83.3% of samples from pigs aged 15-16 weeks, until reaching 97.3% among animals aged > 25 weeks. The overall IgG anti-HEV prevalence in this swine population was 63.6% (Table II). When restricted to animals older than 20 weeks of age, swine IgG anti-HEV was detected in 95.5% of the pigs (107/112). There was a significant difference in the number of IgG anti-HEV seropositive animals when we compared swines from group D (older than 20 weeks of age) with those from group B (3 to 11 weeks of age) and group C (12 to 20 weeks of age) (p < 0.001, 95% CI). This difference was not observed when animals from group D and group A (1 to 2 weeks of age) were compared (p = 0.131, 95% CI). These results suggest that swine HEV infection is largely disseminated (or endemic) within this Brazilian pig population and that the prevalence of IgG anti-HEV antibodies in swines is age-dependent, both characteristics expected for an enterically-transmitted infection in high density farming. Analysis of the seroreactivity to HEV in pigs - Figure shows the distribution of the seroreactivity to HEV according to OD450 values for the several serum samples collected from pigs of different ages when tested by EIA. It could be observed that most of serum samples from animals with ages ranging from 3 to 13 weeks were non-reactive to HEV, showing OD450 values above the cutoff (0.83). On the other hand, most of serum samples from animals in the remaining age groups (1 to 2 weeks and older than 15 weeks of age) were highly reactive to HEV, with OD levels ranging from 0.83 to 3.0. Figure also indicates the mean OD450 values of swine IgG anti-HEV according to age categories (group A to D). There was significant difference in the mean of OD450 values (p < 0.001) when we compare animals from group B (mean OD450 = 0.53 ± SD 0.38) to those from group A (mean OD450 = 1.74 ± SD 0.80), group C (mean OD450 = 1.44 ± SD 0.63) and group D (mean OD450 = 1.78 ± SD 0.57). DISCUSSION Evidence is accumulating that hepatitis E virus infection is a zoonosis (Balayan 1997, Harrison 1999, Meng 2000a,b). This hypothesis has been proposed after the demonstration of substantial seropositivity in different species of animals both in endemic and non-endemic areas, followed by documentation and characterization of swine HEV. The current study presents for the first time data on prevalence rates of HEV-specific antibodies in different animal species and pig handlers in Brazil. HEV seroprevalence studies have been carried out with both commercial and in-house EIAs, using type-common epitopes in ORF2 and ORF3 of the Mexican and Burmese HEV strains as coating antigens (Arankalle et al. 2001, Choi et al. 2003, Hirano et al. 2003). Although these HEV proteins have been developed with the use of human HEV strains, they seemed to be also efficient in detecting anti-HEV antibodies in other animal species (Choi et al. 2003). A recent report demonstrated a 97% concordance in the anti-HEV prevalence results obtained with the human and swine HEV antigens (Eagle et al. 2002), which also supports the presence of cross-species HEV epitopes. In this study, two recombinant proteins were used as antigenic targets in an EIA for the detection of IgG anti-HEV. One protein, pB166, contains the conserved HEV neutralization antigenic epitope, antibodies against which were shown to cross-neutralize different geographic HEV strains (Meng et al. 2001). The other protein, MPII, is composed of strong and broad immunoreactive antigenic epitopes, some of which (ORF3 epitopes) are derived from two different HEV genotypes: the Mexico and Burma HEV strains (Khudyakov et al. 1993, 1994). It was shown that the combination of these two antigens can be used as a very ef-ficient diagnostic target suitable for the reliable detection of HEV-specific antibodies in different epidemiological settings (Obriadina et al. 2002). Our serological data revealed that HEV (or related agent) is highly prevalent in the Brazilian swine commercial herd. In this cross-sectional study of age-prevalence, a dynamic of IgG response to swine HEV similar to that seen in humans infected with enterically-transmitted virus in endemic areas was observed. Of note, 107/112 (95.5%) of the pigs older than 20 weeks were positive for swine IgG anti-HEV, which also showed the highest means of OD450 values. These results show that older animals among this pig population are strongly reactive to swine HEV, suggesting that this virus might be highly disseminated in the environment of this commercial herd. Similar findings were reported in both endemic and nonendemic countries, with seroprevalence rates varying from 15-100% among swine older than 2 months of age (Clayson et al. 1995, Meng et al. 1999, Arankalle et al. 2001, Garkavenko et al. 2001, Takahashi et al. 2003). Several issues raised from the discovery of swine HEV, that seems to be ubiquitous all over the world. The main concern is about the possibility of its transmission to man, since pigs are very common domestic animals and whose meat is also largely consumed. Recently, a small outbreak of hepatitis E was reported in which four people were infected by eating the raw meat of an infected wild deer (Tei et al. 2003). A case control study was further described suggesting that the consumption of uncooked deer meat (in the so-called sashimi form) is an epidemiological risk factor for HEV infection in some areas of Japan (Tei et al. 2004). Indirect evidences also support the hypothesis of zoonotic transmission of HEV to man, such as the high frequency of antibodies to HEV showed by animal handlers (Meng et al. 1999, 2002, Hsieh et al. 1999). In this study, 2 out of 32 pig handlers were IgG anti-HEV positive (6.3%), but this rate was not statistically different from the prevalence observed in normal blood donors (4.3%) from the same geographic region (Trinta et al. 2001). Anyway, further studies are necessary to establish the possible role of swine HEV in human disease. Among the animals investigated in this study, IgG anti-HEV was detected in dogs, cows, chickens and wild rodents, in agreement with other studies (Tien et al. 1997, Favorov et al. 1998, Arankalle et al. 2001). Although a high prevalence has been seen in rats (50%), only four animals were tested, impairing the confiability of this result. However, numerous reports described high anti-HEV prevalence rates in rodents in several geographic regions (Kabrane-Lazizi et al. 1999, Arankalle et al. 2001), and a strain of HEV has been genetically identified from wild rats (Tsarev et al.1998). The high anti-HEV prevalence in rodents suggested that this animal might transmit the virus to swine or vice versa (Meng et al. 2000a,b) since rodents are often found in proximity to swine farms. Macaques (New World monkeys), sheep and goats tested all negative for HEV, in agreement with other sero-prevalence studies that also failed to detected anti-HEV antibodies in some of these animal species (Arankalle et al. 2001, Hirano et al. 2003). In conclusion, our results suggest that HEV is circulating among several animal species, especially swine, in the Southweast Brazil. Further investigations are being conducted to identify the HEV genome in swine with ongoing HEV infection as well as in sewage samples of animal origin for the genetic characterization of the virus. ACKNOWLEDGEMENTS To João Sanches Neto (Emater, Piraí, RJ) for his professional help providing contact with pig farmers from the state of Rio de Janeiro; to Jonaith de Paiva Teixeira Santos for technical assistance in animal care, and Carla Alencar D' Carli, João Carlos Tadeu Streva, and Francisco Louseiro Muniz for authorizing access to their animal facilities during this study. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc05058t1.jpg] [oc05058f1.jpg] [oc05058t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}