 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 100, No. 2, April, 2005, pp. 123-129 Distribution and pathogenicity of Trypanosoma cruzi isolated from peridomestic populations of Triatoma infestans and Triatoma guasayana from rural Western Argentina Marta A Lauricella/+, Raúl L Stariolo*, Adelina R Riarte, Elsa L Segura, Ricardo E Gürtler** Instituto Nacional de Parasitología "Dr. Mario Fatala Chabén"-A.N.L.I.S. "Dr. Carlos G. Malbrán", Av. Paseo Colón 568, 1063 Buenos Aires, Argentina *Coordinación Nacional de Control de Vectores, 5000 Córdoba, Argentina **Laboratorio de Eco-Epidemiología, Facultad de Ciencias Exactas y Naturales, Universidad de Buenos Aires, Buenos Aires, Argentina Financial support: Universidad de Buenos Aires, Ministerio de Salud y Ambiente, Agencia Nacional de Promoción Científica y Técnica, Fundación Mundo Sano, Aventis (ex Agrevo), Chemotecnica (Argentina), NIH Research Grant # R01 TW05836 +Corresponding author. E-mail address: mlauri_2000@yahoo.es Received
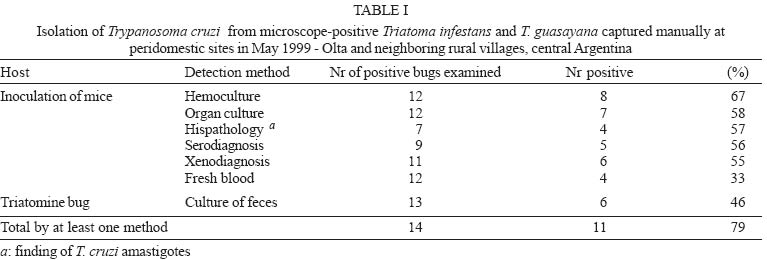
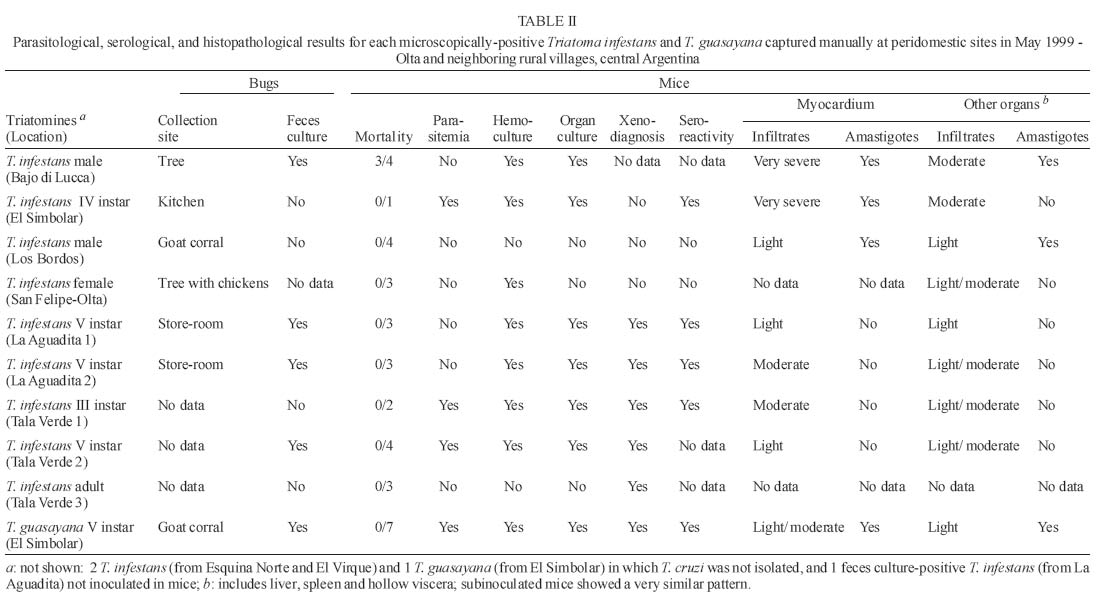
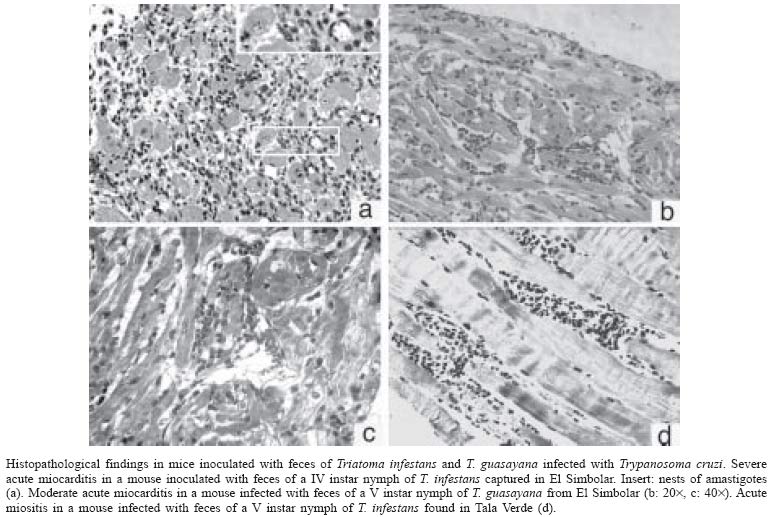
10 November 2004 Code Number: oc05059 We assessed the distribution of Trypanosoma cruzi infection in peridomestic triatomines collected manually at a district-wide scale in rural villages around Olta, Western Argentina, and typed the isolated strains according to their pathogenicity to laboratory mice. Of 1623 triatomines examined, only 14 (0.9%) were infected with T. cruzi based on microscopical examination of feces. The prevalence of T. cruzi infection was 0.8% in Triatoma infestans, 2.3% in T. guasayana, and nil in T. garciabesi, T. platensis, and T. eratyrusiformis. Local transmission occurred in kitchens, store-rooms and goat corrals or nearby, though at very low levels. T. cruzi was detected by at least one parasitological method in 11 (79%) of 14 microscope-positive bugs. Hemoculture was the most sensitive method (67%) followed by culture of organ homogenates, histopathology or xenodiagnosis of inoculated suckling mice (55-58%), and culture of microscope-positive bug feces (46%). The evidence suggests that most of the isolated T. cruzi strains would be myotropic type III. Our study establishes for the first time that peridomestic, microscope-positive T. guasayana nymphs were actually infected with T. cruzi, and may be implicated as a putative secondary vector of T. cruzi in domestic or peridomestic sites. Key words: Trypanosoma cruzi - Triatoma infestans - Triatoma guasayana - Triatoma garciabesi - Chagas disease - surveillance - pathogenicity - Argentina Chagas disease was originally a zoonosis of sylvatic mammals transmitted by triatomine bugs. The process of domestication in Triatominae lead to the simplification of genetic and phenetic characters and may be a generalized current trend (Schofield et al. 1999). Triatoma infestans, probably the main vector of Trypanosoma cruzi, expresses the extreme of an evolutionary trend toward specialization to a stable habitat. But several species of sylvatic or peridomestic triatomines that are not control targets at present might turn into secondary vectors of T. cruzi during or after the elimination of T. infestans (Carcavallo & Martínez 1985). In Northern Argentina, T. sordida, T. guasayana, and T. garciabesi may have potential for domestication in the absence of T. infestans because they have sylvatic or peridomestic colonies and frequently invade human dwellings (Carcavallo & Martínez 1985, Gürtler et al. 1999). The main peridomestic ecotopes of T. gua-sayana are goat or sheep corrals (Canale et al. 2000), but it is usually associated with bromeliads, Opuntia cacti and fallen logs (Wisnivesky-Colli et al. 1997, Noireau et al. 2000). The main peridomestic habitats of T. garciabesi are the rugged bark of Prosopis trees in which chickens roost and coops (Canale et al. 2000). The peridomestic environment includes key sites for the elimination of T. infestans, such as goat or pig corrals and chicken coops (Gürtler et al. 2004), and may be the interface between sylvatic and domestic transmission cycles of T. cruzi, where more parasite diversity can be found. Natural populations of T. cruzi are composed of multiple clones distributed into two major phylogenetic lineages that display distinct biological and eco-epidemiological traits (Tibayrenc 1995, Souto et al. 1996, Barnabé et al. 2000). The prevalence of T. cruzi infection in domiciliary triatomine populations is closely connected to the risk of human infection and depends on human and animal prevalence rates of infection and host contact rates with vectors (Gürtler et al. 1998, Cohen & Gürtler 2001). In rural Northwestern Argentina, a community-wide spraying of deltamethrin caused a sharp fall in T. infestans infection rates with T. cruzi from 49 to 4.6% in domestic sites, and from 6 to 1.8% in peridomestic sites on the 5-year period after spraying (Cecere et al. 1999). The prevalence of T. cruzi infection ranged from 2.4% in T. infestans, 0.7% in T. guasayana, to 0.2% in T. garciabesi, with nearly two-thirds of the infected bugs being caught in perido-mestic sites (Cecere et al. 1999). T. cruzi infections in triatomine bugs are usually diagnosed on morphological grounds in unstained fresh fecal preparations examined by light microscopy. Such infections sometimes were cautiously referred to as "T. cruzi-like trypanosomes" or "flagellates" because light microscopy has limited sensitivity and specificity (Wisnivesky-Colli et al. 1993, Noireau et al. 2000). Other trypanosomatids, such as Blastho-crithidia triatomae (Cerisola et al. 1971) and T. rangeli, may yield "false positive" results. B. triatomae has been found infecting natural populations of Triatominae (e.g. Cecere et al. 1999). As part of a wider project on the effectiveness of pyrethroid insecticides on peridomestic populations of triatomines in Western Argentina (Gürtler et al. 2004), we sought to assess the distribution of T. cruzi infection in peridomestic triatomine populations in an endemic rural district under no vector surveillance; to corroborate that trypanosomes detected by light microscopy were T. cruzi using biological criteria, and to type the isolated strains according to their pathogenicity to laboratory mice. The importance of T. guasayana and other peridomestic triatomines as secondary vectors of T. cruzi was also assessed. MATERIALS AND METHODS Study area - The triatomine bugs were collected in rural areas of the Department General Belgrano and Chamical, Province La Rioja, Argentina. The area, located around Olta (30.4ºS, 66.1ºW), is on a semiarid plain with xerophytic vegetation (Gürtler et al. 2004). Triatomine collection - Six experienced bug collectors searched for triatomine bugs in peridomestic sites of 369 houses in April-May 1999, as described by Gürtler et al. (2004). Timed manual collections in peridomestic sites were conducted using 0.2% tetramethrin for 30 min per house. The collection sites included corrals, pigpens, chicken coops, trees where chickens roosted, piled materials, orchard fences, storerooms, and kitchens. All bugs were examined in the field laboratory to determine numbers by species, stage and collection site as described by Canale et al. (2000), stored and shipped to the laboratory for parasitological examination. Pools of feces from three live or moribund insects of a given species and collection site were examined for flagellate as described. T. cruzi and B. triatomae (Cerisola et al. 1971) were differentiated on a morphological basis. When a positive pool was detected, the feces of each bug were re-examined individually to identify the infected bug for parasite isolation. Isolation methods - In brief, feces from each microscope-positive triatomine were cultured and inoculated into suckling mice which were then examined by fresh blood, hemoculture, culture of organ homogenates, serodiagnosis, xenodiagnosis, and histopathology. Feces culture - Each of the 14 microscope-positive bugs was dissected under sterile conditions to extract the rectal ampoule and prepare an homogenate of feces diluted in 0.4-0.5 ml of Brain Heart Infusion Difco (BHI) (37 g/l). The homogenate was microscopically examined at 400× to determine the number of trypanosomes and then cultured and inoculated into mice. Two scant homogenates were either cultured or inoculated, but not both. Twelve fecal homogenates were inoculated (50-100 µl/tube) into 6 tubes containing 3 ml Nutrient Agar Difco (31 g/l)-0.5 ml defibrinated rabbit blood (Penicillin 200 U/ml-Streptomicin 200 U/ml) as solid slant, and 2 ml BHI-10% fetal calf serum (Bioser, Buenos Aires) as supernatant (Abramo Orrego et al. 1980). Mice studies - Groups of 2-4 C3H suckling mice (15-20 days of age, 8 g) kept in separate cages were inoculated intraperitoneally with 0.1 ml of fecal homogenates (about 1-2 x 103 to 2 x 106 T. cruzi/ml) from each positive bug. Tail blood from each mouse was examined for trypanosomes twice a week. On day 30 post-inoculation, one mouse from each group was studied by xenodiagnosis with 4 T. infestans third instar nymphs during 20-25 min; the pooled feces were examined for T. cruzi 30 and 60 days post-feeding (Cerisola et al. 1974). Hemoculture - Immediately after xenodiagnosis and under anesthesia, heparinized blood was extracted by cardiac puncture and inoculated into 2-3 tubes per mouse without prior centrifugation. On days 45-65 post-inoculation, the remainder mice of each group were processed for sero-diagnosis, hemoculture and histopathology. Organ culture - Heart, liver and spleen samples were homogenated and cultured separately in nutrient agar as before. A total of 12-20 culture tubes from each microscope-positive bug was stored at 28ºC and 50% relative humidity and examined at 400× twice a month for 4-5 months. Small volumes of BHI were added as needed. The supernatant of negative or weakly growing cultures were transferred every 1-2 months into new tubes. Store at liquid nitrogen - Cultured parasites were centrifuged, the pellet dilluted in fetal calf serum-RPMI 80% , ice DMSO- RPMI 80% was added and immediately frozen in liquid nitrogen. Serology - Plasma was tested by an enzyme-linked immunosorbent assay (ELISA) as described previously (Carlomagno et al. 1987). Sera from parasitemic and non infected mice were used as controls. Absorbance values greater than or equal to 0.2 at 490 nm were considered reactive. Anatomopathological studies - Mice inoculated with parasites derived from 7 microscope-positive T. infestans and one T. guasayana were subjected to histopathological studies. Samples of organs were embedded in paraffin and stained with hematoxilin-eosin and Masson's tri-chrome as described by Carlomagno et al. (1987). The intensity of myocardium lesions was graded from 0 (null) to 4 (especially severe acute myocarditis). T. cruzi reinfection in mice - C3H suckling mice were infected with blood trypomastigotes (at least 1 T. cruzi/5 µl blood) of a mouse previously inoculated with feces of T. guasayana. Characterization of strains - Mortality and parasitemia profiles, pathogenicity and tisular tropism were evaluated to characterize the isolates according to Andrade (1974). RESULTS A total of 5251 T. infestans, 379 T. guasayana, 95 T. garciabesi, 6 T. platensis and 2 T. eratyrusiformis was collected from 1748 peridomestic sites inspected. Of the 1623 live or moribund triatomines examined for infection, only 14 (0.9%) bugs were infected with T. cruzi based on microscopical observation of feces. Ten (71%) of the infected triatomines came from different houses and sites, which included trees with or without chickens roosting, store-rooms, kitchens and goat corrals. The percentage of T. cruzi infection was three times larger in T. guasayana (2.3% of 86 bugs) than in T. infestans (0.8% of 1481 bugs) and included 2.3% (2 of 86) and 0.8% (5 of 651) of infected nymphs in each species, respectively. No infected bug was detected among 48 T. garciabesi, 6 T. platensis, and 2 T. eratyrusiformis. T. cruzi was detected by at least one parasitological method in 11 (79%) of 14 microscope-positive T. infestans and T. guasayana (Table I). T. cruzi was isolated more often by hemoculture (67%) than by culture of organ homogenates or xenodiagnosis of inoculated mice (55-58%), or culture of bug feces (46%) or fresh blood examination (33%). A total of 10 (83%) sets of culture tubes (each set originated from one individual bug) remained uncontaminated during the 4-5 month follow-up. T. cruzi was detected by at least one parasitological method in 10 (83%) positive T. infestans, and 8 (67%) isolates were obtained (Table II). Three (27%) groups of mice only showed very low parasitemia (less than 5 parasites per 5 µl of blood), and other three were parasitemia-negative and xenodiagnosis-positive. Four (67%) groups of mice tested by ELISA between 23 and 65 days post-inoculation were seroreactive for T. cruzi at serum dilutions from 50 to 1600. All of the seroreactive mice were organ-, hemo-culture- or histopathology-positive for T. cruzi. In the two groups of seronegative mice, the Los Bordos strain was amastigote-positive by histopathology whereas the San Felipe-Olta strain was hemoculture-positive. In general, histopathology of the T. cruzi strains isolated from T. infestans revealed myocardium and skeletal muscle involvement at 1-2 months post-inoculation. Low mortality at the first month, low parasitemia and virulence suggest that these strains belonged to type III. The Bajo di Lucca strain induced moderate to very severe pathologic changes in all organs, and was the only one to produce heavy mortality at 28-30 days post-inoculation. The myocardium had a very severe and intense inflammatory process, associated to a moderate number of amastigote nests. The liver had a moderate inflammation with no amastigotes found, whereas hollow viscera showed slight inflammation with scarce amastigote nests. El Simbolar strain induced very severe myocarditis, with numerous amastigote nests (grade 4) (Fig. 1a); moderate inflammation in the liver but not lesions in hollow visceras. The Los Bordos strain induced a light inflammatory infiltration, with amastigote nests only in heart and striated muscle. The La Aguadita and Tala Verde strains 1-2 induced light to moderate inflammatory infiltrations but no amastigotes were seen (Table II, Fig. 1d). T. cruzi was recovered from only one of the two positive T. guasayana (Table II). Mice inoculated with feces from this T. guasayana had low levels of parasitemia (< 4 parasites per slide) during most of the follow-up and positive xenodiagnosis at 30 days post-inoculation. Both the inoculated and reinoculated mice were seroreactive for T. cruzi. The T. guasayana-derived strain from El Simbolar induced a light to moderate cardiac inflammatory process (Fig. 1b,c), with some amastigote nests. Striated muscle, liver and hollow viscera presented a light or moderate diffuse inflammatory process, but amastigotes were either scarce or absent. Mice reinoculated with this strain showed a very similar parasitologic and histophatologic pattern as the inoculated mice. DISCUSSION Our study shows that peridomestic triatomines from rural Western Argentina were actually infected with mostly myotropic T. cruzi strains, though at very low infection rates, and peridomestic T. guasayana was three times more infected with T. cruzi than T. infestans. Most (83%) of the microscopically detected trypanosomes proved to be T. cruzi according to standard biological criteria (Hoare 1972). In addition to being morphologically indistinguishable from T. cruzi, the trypanosomes were successfully cultured in specific media, induced a detectable serological response to T. cruzi, and were pathogenic to laboratory mice. The infected mice were also infective to T. infestans and reinoculated mice, allowing the continuity of infection. To our knowledge, this may be the first study that applies these criteria to establish that positive peridomestic triatomines, particularly T. guasayana, were actually infected with T. cruzi. Parasites were isolated from two-thirds of the infected bugs. Isolation of T. cruzi from bug feces has not been favored in the past because of fungal and bacterial contaminations, but here only two of 13 cultures of T. infestans feces presented fungal contaminations. In both cases, all isolation attempts were negative but histopathological studies revealed T. cruzi amastigotes and light lesions. In addition, seroreactivity for T. cruzi failed twice to reveal mouse infections in which parasites were subsequently isolated. Mice infected with Los Bordos strain were not seroreactive at 65 days post-inoculation though histopathological findings were positive. Suckling mice inoculated with T. infestans and T. guasayana feces displayed serological, parasitological, and pathological findings compatible with T. cruzi. All strains isolated from T. infestans and the one from T. guasayana apparently were myotropic type III, with slow parasite multiplication, low virulence, no mortality during the first month of infection, and pathogenicity especially affecting skeletal muscles and myocardium (Andrade 1974, Andrade 2000). Moderate lesions and scarce amastigote nests were detected in mice inoculated with T. guasayana-derived T. cruzi, whereas more severe lesions were produced by some strains isolated from T. infestans. Most mice inoculated with feces from T. infestans and T. guasayana developed very low parasitemia and suffered low mortality, probably mostly caused by low infecting doses, as the field-collected triatomines usually had a low intensity of infection on arrival to the laboratory. Mice infected with T. guasayana-derived parasites showed a pattern similar to acute Chagas disease. Among all the trypanosomatids detected in humans or mammals in the Americas, T. cruzi is the only agent causing tissue pathology. T. rangeli is not pathogenic to humans, and so far has not been detected in Argentina (D'Alessandro & Del Prado 1977). The different methods herein used confirm that the trypanosomatids isolated from T. guasayana were T. cruzi beyond any doubt. Our district-wide study confirms that in rural semiarid Argentina, peridomestic triatomine populations have very low rates of T. cruzi infection. Microscopical examination of triatomine feces may have underestimated the true bug infection rates to some extent, as suggested by paired comparisons with the polymerase chain reaction (Breniere et al. 1995). Interestingly, peridomestic T. guasayana had a non significative three times larger infection rate than T. infestans despite being less abundant. T. guasayana has been considered a potential risk for humans because of its widespread geographic range and larger capacity to endure low temperatures than T. infestans (Carcavallo & Martinez 1985). More recently in Northwestern Argentina, T. guasayana frequently invaded homes and bit people (Gürtler et al. 1999); it was found infected with T. cruzi (Cecere et al. 1999) and significantly associated with transmission to native dogs during the surveillance phase (Castañera et al. 1998). T. guasayana thus appears implicated as a putative secondary vector of T. cruzi in domestic or peridomestic sites under certain eco-epidemiological scenarios. Some of the peridomestic triatomines infected with T. cruzi were collected in kitchens or store-rooms. As these are resting sites of dogs or cats and both are highly infective to bugs (Gürtler et al. 1998), these were the likely sources of infections. However, other infected adults and nymphs were captured from goat or pig corrals and chicken coops, but goats and pigs only exceptionally have been infected with T. cruzi (Pinto Dias 2000) and chickens are refractory to infection. Light-trapping collections of large instars of T. infestans and T. guasayana suggested active walking dispersal from potential sources located about 50 m away (Vazquez-Prokopec et al. 2004). Therefore, the finding of two infected nymphs of T. guasayana in goat corrals located 10-40 m away from bedrooms suggests that local transmission occurred there or nearby, though at very low levels and from as yet unidentified sources. Ongoing work is directed to establish whether these infections are linked to the domestic or sylvatic transmission cycle of T. cruzi by means of molecular typing techniques. ACKNOWLEDGEMENTS To Fundación Mundo Sano, Aventis (ex Agrevo) and Chemotecnica (Argentina) for financial, administrative and technical support. A.N.L.I.S-Ministerio de Salud y Ambiente, The University of Buenos Aires and Agencia Nacional de Promoción Científica y Técnica gave partial support to this project. To Roberto Chuit, Rodrigo González Llanos, Rubén Di Lello, Delmi Canale, Daniel Salomón, Cynthia Spillmann, and the ECLAT network for their support. REG and ELS are members of the Researcher's Career from the National Council of Scientific and Technologic Investigations (Conicet) from Argentina. The final stage of the study was supported by NIH Research Grant # R01 TW05836 funded by the Fogarty International Center and the National Institute of Environmental Health Sciences (NIEHS)(Uriel Kitron and REG, co-PI). REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc05059t1.jpg] [oc05059t2.jpg] [oc05059f1a-d.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}