 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
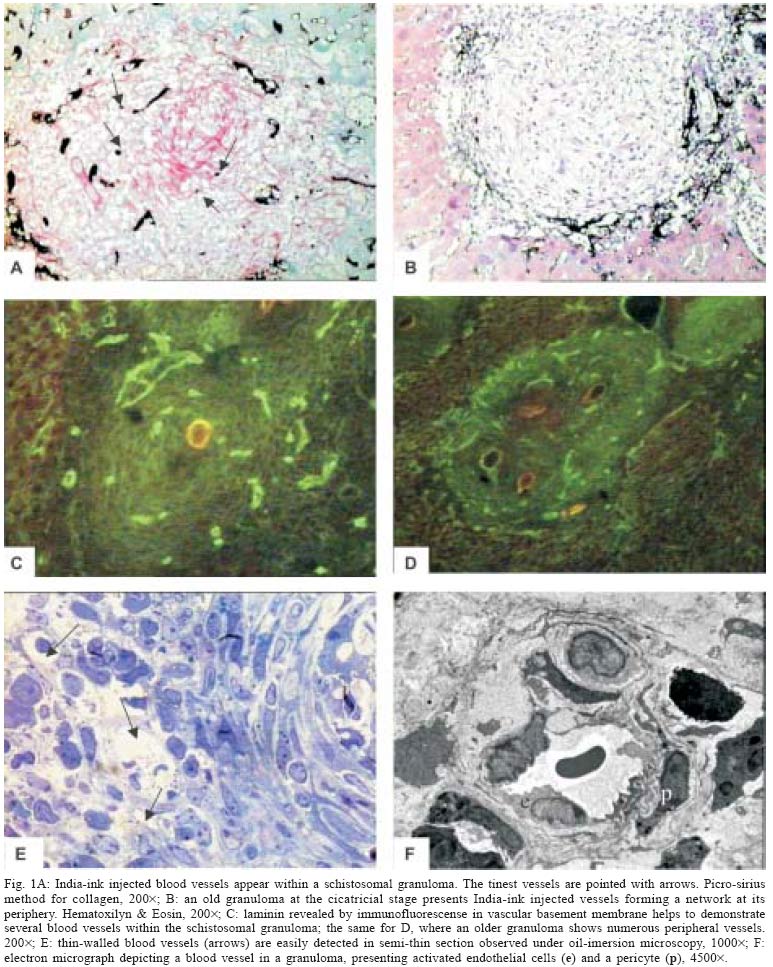
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 100, No. 2, April, 2005, pp. 183-185 SHORT COMMUNICATION Angiogenesis and schistosomal granuloma formation Ana Paula Baptista, Zilton A Andrade+ Laboratório de Patologia Experimental, Centro de Pesquisas Gonçalo Moniz-Fiocruz, Rua Valdemar Falcão 121, 40295-001 Salvador, BA, Brasil Financial
support: Papes III, Pibic-Fiocruz Received
25 November 2004 Code Number: oc05067 An increasing amount of evidences points to angiogenesis as playing a paramount role in fibrosis development. However, granulomas in general, and periovular schistosomal granulomas in particular, are considered avascular structures, although they usually result in dense areas of focal fibrosis. In order to clarify this apparent paradox, the presence of blood vessels was systematically searched in hepatic schistosomal granulomas of mice, during different stages of the infection, and at different stages of granuloma evolution, by means of vascular injections of colored masses, demonstration of laminin in vascular basement membranes and by ultra structural analysis. Vascular proliferation appeared evident at the early stages of granuloma formation, gradually decreasing thereafter, older granulomas becoming almost avascular structures, sometimes delimited at the periphery by a rich vascular network. Key words: Schistosoma mansoni - periovular granuloma - angiogenesis. Schistosoma mansoni, one of the blood flukes, lives within the host vascular system. The eggs laid down by the female worms are carried out by the portal blood flow until they stop within hepatic pre-capillary venules when the diameter of the vessels does not permit further progression. Thus, they are impacted and acquire an intimate contact with endothelial cells, before rupturing the vessel walls to reach the interstitial tissues. Lenzi et al. (1988) demonstrated endothelial cells, immunohistochemically identified by factor-VIII, present in great number in schistosomal periovular granulomas. They suggested that endothelial cells play a fundamental role, not only in granuloma formation, but in the pathogenesis of schistosomiasis in general. Additionally, studies in vitro have revealed that soluble egg antigens (SEA) induce proliferation of endothelial cells (Freedman & Ottesen 1988), and also up-regulate vascular endothelial growth factor and angiogenesis (Loeffler et al. 2002). However, in a classical textbook (Florey 1954), granulomas were considered structures devoid of blood vessels. One study that particularly explored the vascular lesions in experimental schistosomiasis of mice, depicted the granulomas in the liver as avascular structures, surrounded by numerous India-ink injected vessels (Cheever 1961). As a matter of fact, little is known about the in situ kinetic relationship of blood vessels and the parasite eggs within the periovular granulomas. Since angiogenesis appears to be a key factor for new connective tissue synthesis, an investigation on the behavior of blood vessels during schistosomal granuloma formation seemed appropriated. Fragments of the liver were taken from adult mice during early (8 weeks from exposure) and late (16 to 20 weeks) S. mansoni infection. Infection was made by the transcutaneous route, with 50 freshly eliminated cercariae. The liver tissues were submitted to the following procedures: (1) neutral formalin fixation and paraffin embedding, followed by staining of the sections with hematoxilyn-eosin, sirus-red for collagen, and orcein for elastic fibers; (2) other fragments were immediately frozen in liquid nitrogen and submitted to cryostat sectioning for the application of the immunofluorescence technique with a rabbit anti-human laminin to be revealed with fluoresceinated anti-rabbit IgG and microscopically examined under UV-light; (3) tiny pieces of tissue were fixed in 4% glutaraldehyde in 0.2M cacodylate buffer, pH 7.4, and postfixed with 1% osmium tetroxide and embedded in Polybed. Ultra-thin sections were contrasted with uranyl acetate and a lead citrate and examined in a Zeiss EM-9 electron microscope, at 50 kV. On another set of experiments, animals with either recent and old infections were anesthetized and submitted to a direct manual injection of the portal system with a mixture of 33% India-ink and 12% gelatin, until a uniform blackening of the liver surface was obtained. After injection the liver was removed and fixed in 10% formalin. Slices were embedded in paraffin and the 10-20 µ-thick sections were lighly stainined with hematoxylin and microscopically examined. Blood vessels were difficult to be observed in routinely stained histologic sections, even when orcein stain for elastic fibers was used. However, well vascularized periovular granulomas were easily detected in preparations made with India-ink injection (Fig. 1A) and by immunofluorescent laminin staining of the vascular basement membranes (Fig. 1C) in both early and late infection, more frequently in the former. Some granulomas exhibited none or only a few injected vessels. These granulomas were more fibrotic, centered by desintegrated eggs or simply by egg shells, and usually presented injected blood vessels at their pariphery, sometimes forming a profuse network (Figs. 1B, 1D). The vessels within the granulomas were usually tiny capillaries that probably are missed during routine histologic examination. When thin resin-embedded sections are examined by oil-imersion (Fig. 1E) or ultra-thin sections by electron microscopy (Fig. 1F), these vessels are more frequently detected. However, the presence of such vessels is variable according to the stage of development of the granuloma, both in early and late infections. Vessels were more frequently detected within the granulomas during early as opposed to late infection. This was related to the increase in numbers of more mature or fibrotic granulomas, as the infection lasted longer. Fibrotic granulomas showing remnants of egg shell in their center tended to exhibit a vascular network at the peri-phery. From the point of view of general pathology periovular schistosomal granulomas are dynamically similar to wound healing, with production of granulation tissue, which becomes less and less vascularized with time, while the cicatrial fibrous tissue becomes more compact and mature. This association of connective tissue neo-formation and differentiation with vascular proliferation is well represented during schistosomal granuloma formation. A growing number of papers are pointing out to the role of angiogenesis in the genesis and progression of hepatic fibrosis and cirrhosis. A recent study associates ischemia, production of pro-angiogenic factors, such as vasculo-endothelial growth factor and vascular proliferation in the generation of fibrosis due to experimental bile duct ligation (Ramarduc et al. 2003) An investigation on liver fibrosis associated with Capillaria hepatica infection in rats disclosed that angiogenesis preceded collagen deposition when very early septa were just starting to sprout from portal spaces (Souza 2003). Angiogenesis inhibitors are being indicated as an effective tool for the therapy of hepatic fibrosis (Croquete et al. 2003). In conclusion, the general morphologic pattern of fibrosis formation during periovular schistosomal granuloma is in keeping with the concept that angiogenesis is at the roots of fibrogenesis. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz. The following images related to this document are available:Photo images[oc05067f1a-f.jpg] |
| |||||||||

{kind=link}