 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
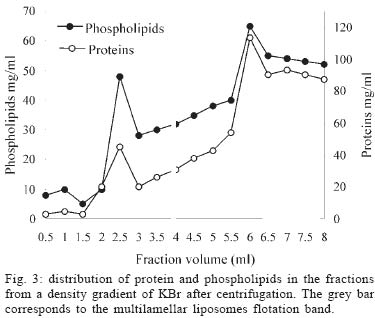
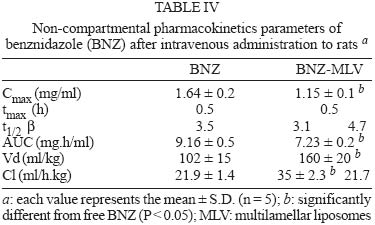
Mem Inst Oswaldo Cruz, Rio de Janeiro, Vol. 100, No. 2, April, 2005, pp. 213-219 Benznidazole vs benznidazole in multilamellar liposomes: how different they interact with blood components? Maria Jose Morilla++, Maria Jimena Prieto, Eder Lilia Romero+ Laboratorio de Diseño de Estrategias de Targeting de Drogas, Departamento de Ciencia y Tecnología, Universidad Nacional de Quilmes, Roque Saenz Peña 180, Bernal, 1876, Buenos Aires, Argentina Financial
support: Secretaria de Investigaciones de la UNQ. Received
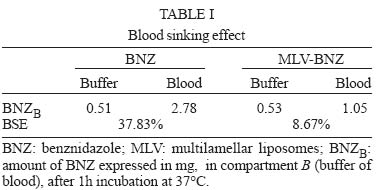
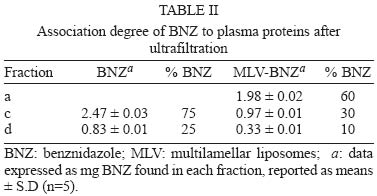
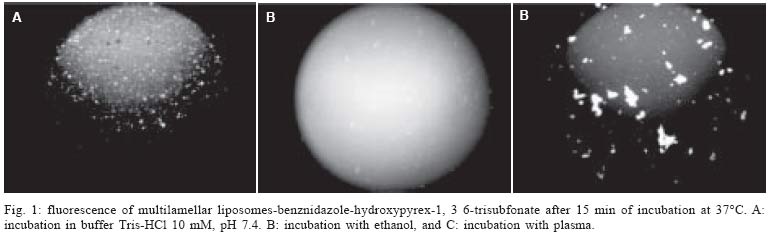
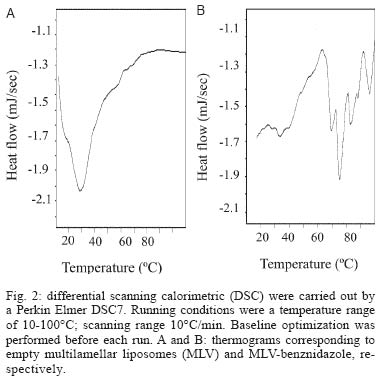
12 August 2004 In spite of its widespread use, benznidazole's (BNZ) toxicity and low efficacy remains as major drawbacks that impair successful treatments against Chagas disease. Previously, attempting to increase the selectivity and reduce its toxicity on infected tissues, multilamellar liposomes (MLV) composed of hydrogenated soybean phosphatidylcholine (HSPC): distearoyl-phosphatidylglycerol (DSPG): cholesterol (CHOL) 2:1:2 mol:mol loaded with BNZ (MLV-BNZ) were designed. In this work we compared different properties of MLV-BNZ with those of BNZ. Opposite to other hydrophobic drugs, the results indicated that slight changes of BNZ´s association degree to proteins and lipoproteins should not modify the percentage of unbound drug available to exert pharmacological action. On the other hand, when loaded in MLV, BNZ reduced its association to plasma proteins in 45% and became refractory to the sinking effect of blood, dropping 4.5 folds. Additionally, when loaded in MLV, BNZ had higher volume distribution (160 ± 20 vs 102 ± 15 ml/kg) and total clearance (35.23 ± 2.3 vs 21.9 ± 1.4 ml/h.kg), and lower concentration-time curve (7.23 ± 0.2 vs 9.16 ± 0.5 µg.h/ml) than BNZ. Hence, these studies showed that for MLV-BNZ, the amount of BNZ can be substantially increased, from 25 to 70%, being this formulation more rapidly cleared from circulation than free drug; also due to the lower interaction with blood components, lower side effects can be expected. Key words: liposomes - benznidazole - Chagas disease Trypanosoma cruzi, the causative agent of Chagas disease, is a parasite that follows a complex cycle inside the mammalian host, alternating extra and intracellular stages. The extensive binding to plasma proteins expected for the hydrophobic benznidazode (BNZ), the only antichagasic drug currently approved in Argentina and Brazil (Coura & de Castro 2002), is in part responsible for its distribution in almost all types of tissues. This fact makes difficult the targeting of therapeutic amounts of BNZ to infected cells. Liposomes are potential candidates for increasing the selective delivery of this drug since they could be captured by infected tissues. In a previous work by our group, BNZ has been loaded in multilamellar liposomes (MLV-BNZ) and intravenously (iv) injected, aiming to direct its biodistribution to the liver (Morilla et al. 2002). Administration by iv of a single bolus of MLV-BNZ three times a week during two weeks led to an increased selectivity for the liver with an increased accumulation of BNZ in this organ, when compared with the same dose of the drug itself (Morilla et al. 2004). However, to fully exploit the differences between BNZ and MLV-BNZ, pharmacokinetics data are still required. When liposomal drugs are iv injected, a complex set of phenomena occurs, starting with the rapid adsorption of plasma proteins on the liposome surface. Two types of proteins play a major role in this interaction: opsonins, specialized in tagging the liposomal surface to make it recognizable to be phagocytozed by Kupffer cells (KC) in the liver (Patel 1992, Kulkarni et al. 1995, Hiroshi 1997); and lipoproteins (Maurer et al. 2001). In general, the adsorption of plasma proteins has no other physical consequences than promoting the uptake of particles by KC, but that is not the case for the interaction of liposomes and lipoproteins. Whereas LDL mainly possesses the integral and not exchangeable apolipoprotein B, HDL possesses the hydrosoluble and exchangeable apolipopro-teins A, C and E and only the interaction between liposomal membrane and apolipoprotein leads to bilayer disruption (Rodrigueza et al. 1998). In this case, hydrophilic drugs leak from the aqueous phase of the liposome. An hydrophobic drug such as BNZ, dissolves in phospholipid bilayers according to a partition coefficient (Kp = [drug]liposomal bilayer/ [drug]aqueous medium); therefore, since liposomes generally suffer up to a 500-2000 folds dilution when injected by iv, an efflux of drug towards the external medium follows their administration. Therefore, upon iv injecting MLV-BNZ, two fractions are expected: a, liposomal BNZ (retained in liposomal matrix), and b, released BNZ. Fraction a can not escape from blood circulation and is mostly phagocyted by KC and other accessible macrophages; this process is considered a non-conventional biodistribution of the drug. On the other hand, the interaction between fraction b and blood leads also to two drug fractions: c, bound BNZ (BNZ associated to proteins and/or blood cells) and d, unbound BNZ. This latter fraction could cross the endothelial lining of the blood vessels, get into tissues leading to pharmacological action following a conventional biodistribu-tion, opposite to fraction c which remains in the vascular compartment as an inactive pool. In this work we compared the association of BNZ to blood with that of MLV-BNZ, specifically to plasma proteins and lipoproteins, i.e. fractions c and d. In case of MLV-BNZ, we also quantified fractions a and b. In the literature there is a scarcity of studies about the association of BNZ and of liposomal BNZ with total plasma proteins, lipoprotein fraction, and blood cells. The knowledge of such interactions as well as their role in the pharmacokinetics characteristics of both formulations will contribute to gain insights on their expected trypanocidal and toxic effect in vivo. MATERIALS AND METHODS Chemicals - BNZ (purity: 99.85%) was supplied by Roche Argentina (RO-07-1051/000). Hydrogenated soybean phosphatidylcholine (HSPC) and distearoyl-phosphatidylglycerol (DSPG) were from Northern Lipids (Vancouver, Canada). Cholesterol (Chol) and KBr were purchased from Sigma (St Louis, US). Dimethyl sulfoxide (DMSO) and acetonitrile (ACN) were of HPLC-quality (Carlo Erba Reagenti, Italy). Tris, trichloroacetic acid (TCA), chloroform and methanol were of analysis grade (Anedra, Argentina). The fluorophore 8-hydroxypyrene-1, 3, 6-trisulfonate (HPTS) and the quencher p-xylene-bispyridinium bromide (DPX) were purchased from Molecular Probes (OR, US). Normal human plasma was obtained from plasma bank. Liposomal preparation - Briefly, MLV-BNZ composed of HSPC:DSPG:Chol 2:1:2 mol:mol were prepared as described by Morilla et al. (2002), by mixing the lipids from solutions in Cl3CH: CH3OH (9:1, v/v) and BNZ in DMSO at 5/100 (mol/mol) drug to lipid ratio. The solvent was evaporated in a rotoevaporator at 50°C. The resulting film was flushed with N2, and suspended in 500 µl of 10 mM Tris-HCl buffer (pH 7.4) to a final concentration of about 20-30 µmol/ml total lipids. The BNZ non-incorporated to the liposomal matrix was eliminated by centrifugation at 6000 g for 10 min and the liposomal pellet was resuspended in Tris-HCl buffer. Lipid concentration was determined by a colorimetric phosphate micro assay (Bötcher et al. 1961). The encapsulated BNZ was determined after complete disruption of a liposomal sample in DMSO (1:10, v/v) and quantified by HPLC coupled to UV in a Beckman System Gold with a Programmable Solvent Module 125 and a Programmable Detector Module 166 (Morilla et al. 2003). Quantitation was based on the peak area-concentration response of the HPLC calibration curves. The wavelength was set at 324 nm with a response time of 1 s. The analytical column was a reverse phase Kromasil C18 (25 × 0.4 cm i.d., 100 Å, 5 µm particle size). The mobile phase used was ACN-water (40:60, v/v), degassed by vacuum. Elution was performed isocratically at a flow rate of 0.9 ml/min. Blood sinking effect - We used a plastic chamber which consisted of compartment S (sample), containing 2.5 ml BNZ or MLV-BNZ (6 µg BNZ) in 10 mM Tris-HCl pH 7.4, separated by a polycarbonate membrane (200 nm pore size) from compartment B (blood), containing 5 ml buffer or blood. Free or released BNZ can diffuse through the membrane from S to B, while liposomes, due to their size (nearly 1-3 µm) are retained (Morilla et al. 2004). Before and after incubation under agitation for 1 h at 37°C, BNZ in compartment S was quantified as described in the previous item. When buffer was in compartment B, BNZ was quantified as described above, and in the case of blood the drug was extracted by addition of 400 µl ACN-DMSO (1:1, v/v) to 200 µl blood, followed by incubation at 4°C for 30 min. After centrifugation at 6000 g for 20 min, the supernatant was incubated with 100 µl TCA solution 10% (w/v) for 15 min at -12°C and centrifuged for 10 min at 10000 g. Finally, 20 µl from this latter supernatant was quantified by HPLC as described above. Blood sinking effect (BSE) was calculated as the percentage of BNZ captured in compartment B filled with blood minus that captured in compartment B filled with buffer (corresponding to only diffusion of the drug, without active capture). Association degree of BNZ to plasma proteins - After incubation BNZ or MLV-BNZ (6.6 µg BNZ) with 2 ml of plasma for 15 min at 37°C, 1 ml of the mixture was ultrafiltrated by centrifugation for 1 h at 5000 g in an Ultrafree-CL PTMK polysulfone filter unit, a long-lived highly protein-retentive membrane of 300,000 nominal molecular weight limit (Millipore Corporation Bedford, US). Afterwards BNZ was quantified by HPLC both in fractions c, (non filtered BNZ; upper compartment) and d, (filtered unbound BNZ; lower compartment). In case of MLV-BNZ, the whole content of the upper compartment was centrifuged for 20 min at 6000 g to separate fraction a (pellet BNZ retained in liposomal matrix) from fraction c, (supernatant, bound BNZ). The drug was quantified in both fractions. The BNZ mass/plasma volume ratio used in the assays was 3.3 µg BNZ/ml plasma, resulting in 40-fold dilution of MLV-BNZ and corresponding to the concentration of MLV-BNZ injected by iv in rats (Morilla et al. 2004). As control plasma alone was submitted to ultrafiltration and proteins in the upper and lower compartments were quantified as previously described (Bradford 1976), in order to determine the amount of low molecular weight proteins capable of passing the filter. A mixture of plasma with MLV loaded with the fluorescent dye HPTS (see next item) was also ultrafiltrated, to check for the maintenance of liposomal structural integrity, determined by dye leakage and phospholipid quantitation in the lower compartment. Morphological and permeability changes on liposomes induced by plasma - MLV-BNZ were prepared as described above except that 35 mM HPTS and 50 mM DPX were added to 10 mM Tris-HCl buffer (pH 7.4). The non-encapsulated HPTS and DPX were eliminated by one step of centrifugation and the liposomal pellet was re-suspended in the same buffer. The fluorescent labelled liposomes, MLV-BNZ-HPTS, were incubated 15 min with plasma at 37°C and the emission of HPTS at 510 nm resulted from excitation with blue filter (450-490 nm) was seen with in an Olympus BH2-RFGA fluorescence microscope Association degree of BNZ to lipoproteins - Lipoprotein separation was performed as described by Cassidy et al. (1998). Briefly, after incubation of plasma with BNZ or MLV-BNZ for 15 min at 37°C, the density of the system was increased to 1.25 g/ml by addition of solid KBr and 3 ml of the mixture was placed on the bottom of an Ultraclear centrifuge tube (Beckman Instruments, CA, US) and 2.8 ml of three KBr solutions of different densities (δ = 1.21, 1.063 and 1.006 g/ml) were carefully layered on it. The tubes were ultracentrifuged on a SW 40 Ti swinging bucket rotor at 40,000 rpm for 18 h at 15°C in a Beckman XL-70 ultracentrifuge. Afterward, fractions of 0.5 ml were collected from top to bottom of the tubes for quantification of protein (Bradford 1976), phospholipid (Bötcher et al. 1961) and BNZ. Pharmacokinetics parameters - Wistar rats (180-250 g body weight) received a single dose of 0.2 mg/kg BNZ in Tris-HCl pH 7.4 or MLV- BNZ in the lateral tail vein. Animals were divided into two groups. The first one (n = 5) was bled at 0.5, 1, 2 and 3 h post injection (pi) and sacrificed at 4h pi. And the second (n = 5) at 5, 6, 7 and 8 h pi and sacrificed at 9 h pi. Blood samples were collected from the retroorbital sinus on tubes containing sodium citrate and BNZ was quantified by HPLC. The non-compartamental pharmacokinetic parameters: area under the concentration-time curve (AUC), total clearance (CL), apparent volume of distribution (Vd) and biological half-life (t1/2) were calculated based on the method described by Gibson and Perrier (1982). The data were analyzed for statistical significance by the t-test (P < 0.05) and the results were expressed as the mean ± standard deviation (S.D.). RESULTS AND DISCUSSION Blood sinking effect - In order to test the capacity of blood components of acting as a sink of BNZ, we designed a simple chamber based on the unidirectional efflux of BNZ from compartment S to B across the filter. Using this chamber, direct contact between blood and liposomes was avoided, as described, for instance, by Hinderling (1997). The results shown in Table I indicated that blood sinking effect (BSE) was higher for BNZ (37.83%) than for MLV-BNZ (8.67%). For other infectious diseases such as malaria, several beneficial effects of drug partitioning in erythrocytes have been documented (Cenni & Betschart 1995, Charache et al. 1995, Snoek et al. 1996, Venitz et al. 1987), but also toxic effects resulted from those interaction, such as increased formation of methemoglobin and hemolysis (Grossman & Jollow 1988, Fasanmade & Jusko 1995). Since BNZ acts on circulating trypomastigotes and on intracellular amastigotes of T. cruzi, which do not colonize erythrocytes, its partitioning on blood cells should be considered potentially toxic; hence, the reduced interaction of BNZ when loaded in MLV could be beneficial. Association degree to plasma proteins - Equilibrium dialysis and ultrafiltration are the most used methods to provide both accurate and precise assessment of drug's binding to plasma protein resembling the in vivo binding situation (Koike et al. 1985). By ultrafiltration of plasma alone, no proteins were detected in the lower compartment, ensuring that only unbound BNZ was quantified, when mixtures of BNZ and plasma were assayed. No filter stacking by vesicles neither liposomal leakage from the upper compartment were determined by visual inspection of the dye after ultrafiltration of a mixture of MLV-BNZ-HPTS and plasma. The amounts of BNZ found in fractions a, c, and d after ultrafiltration are shown in Table II. Ultrafiltration of BNZ plus plasma resulted in a percentage of BNZ of 25 ± 0.5% in fraction d and of 75 ± 1% in fraction c. The corresponding values for MLV-BNZ plus plasma were of 10 ± 0.2% in fraction d and 90 ± 0.5% in the upper compartment. The content of the upper compartment was centrifuged to separate the MLV-BNZ from plasma, and this 90% resulted distributed 60 ± 0.5% in fraction a and 30 ± 0.2% BNZ in fraction c. This percentage of 60% retained in liposomal matrix, together with the release of the drug of about 40% upon a 40-fold dilution, were coincident with those of previous experiments in which MLV-BNZ was diluted in buffer (Morilla et al. 2002). Interestingly, the ratio bound/unbound drug (3/1) was the same both for BNZ and MLV-BNZ. In sum, the ultrafiltration experiment indicated that when 0.2 mg BNZ as MLV-BNZ/kg body weight was injected by iv, only 40% of total BNZ was available to act as free drug. In this context the association to plasma proteins was reduced from 75 ± 1% in BNZ to 30 ± 0.2% in MLV-BNZ, and in the case of the liposomal formulation unbound BNZ was reduced from 25 ± 0.5% to 10 ± 0.2%. These results taken together show that for MLV-BNZ 70% of the drug was available to exert its pharmacological action, being 10% conventional and 60% non-conventional biodistribution, against 25% in the case of BNZ alone. Morphological and permeability changes on liposomes induced by plasma - Screening of dynamic changes of liposomes in plasma, such as aggregation and leakage of liposomal aqueous content, was done using MLV-BNZ-HPTS and plasma. HPTS is a hydrosoluble fluorescent dye, which when co-encapsulated with the collisional quencher DPX, presents a low intensity emission at λem= 510 nm after excitation at λexc = 440-490 nm. This indicates that HPTS remains trapped in the inner aqueous phase and therefore the liposomal integrity is maintained (Daleke et al. 1990, Straubinger et al. 1990, Yoshimura 1995). Fig. 1A shows the typical spherical shape with confinement of the fluorescent dye to the inner aqueous phase of the MLV, and in Fig. 1B the dye leakage (uniform fluorescence) upon disrupting the vesicles in ethanol is shown. In the presence of plasma, liposomes aggregated, loosing their spherical shape without significant leakage (Fig. 1C). Maintenance of membrane integrity was probably due to the low interaction with lipoproteins (in the order of 4-5%, see next item). On the other hand, differential scanning calorimetric (DSC) measurements of empty MLV (liposomal matrix) showed a main phase transition temperature at 29°C (Fig. 2A), whereas MLV-BNZ showed a main phase transition temperature at 69-75°C (Fig. 2B). Therefore, the temperature of 37°C used in the experiment was bellow that of phase transition, indicating that the membranes are in the gel phase, a state in which bilayers are refractory to protein adsorption. Association degree of BNZ to lipoproteins - When liposomes and HDL are mixed together, the outcome of their mutual interaction depends on their relative amounts. Since the mass of liposomal phospholipid used in this work was 22 mg/kg body weight, and the solubilizing capacity of HDL is only exceeded beyond 20-50 mg phospholipids/kg body weight (Scherphof et al. 1978, Williams et al. 1984, Willians & Scanu 1986, Huuskonen et al. 1996, Lusa et al. 1996), liposomes were exposed to the maximal solubilizing capacity of HDL apolipoproteins in all the experiments. The protein and phospholipid content of each 0.5 ml fraction resultant from plasma ultracentrifugation is shown in Fig. 3. The fractions corresponding to 1 ml, 2.5 ml and 6 ml were ascribed to VLDL, LDL and HDL since their protein/phospholipid ratio were coincident with those of each lipoprotein. MLV loaded with HPTS floated as a well defined green band, isolated from the lipoprotein bands. The association degree of BNZ or released BNZ to lipoproteins, determined as amount of BNZ extracted from each lipoprotein fraction, is shown in Table III. The results indicated that only 5% of BNZ associated to lipoproteins, distributed 4.2 ± 1% in HDL and 0.8 ± 0.6% in VLDL and that 4% of MLV-BNZ associated to lipoproteins, distributed 2.8 ± 0.9% in HDL, 0.5 ± 0.2% in VLDL and 0.5 ± 0.2% in LDL. These findings indicated that, independently of the intrinsic differences, the association degree of BNZ to lipoproteins, as BNZ or MLV-BNZ, was fairly below those of hydrophobic drugs such as nystatine, amphotericin B and A cyclosporine, free or as liposomal forms, which was situated between 50 to 70% (Awni & Sawchuk 1985, Wasan et al. 1993). Since only high association degrees had been shown to modify the pharmacological activity of a drug (Awni & Sawchuk 1985, Wasan et al. 1993, 1997a), we can expect that the association degree of 4-5% between BNZ or MLV-BNZ and lipoproteins have no impact on their behaviour as drug effect. Besides, anionic phospholipids derived from phosphatidyl-glycerol-containing liposomes act as bridges between HDL and a hydrophobic drug promoting the transfer from the liposomal form drugs to HDL and consequently modifying its distribution pattern (Surewiicz et al. 1986, Wasan et al. 1993). For instance, the incorporation of nystatin, amphotericin B or A cyclosporine into phosphatidyl- glycerol containing liposomes increased drug association to HDL and lowered their association to LDL as compared to non liposomal drugs; consequently, their inherent toxicity is reduced (Wasan et al. 1997b, Cassidy et al. 1998). However, in the presence case the DSPG-containing liposomal matrix of MLV-BNZ did not increase the transference of BNZ to HDL above the association degree of BNZ. Many infectious processes are associated with severe dislipemia and changes in relative amounts of lipoproteins. Previous studies have shown that asymptomatic chagasic patients present a lower HDL content than healthy individuals (Cano et al. 1985). If the extent of BNZ association to lipoproteins was high, it could be expected that the reduced levels of HDL could influence the biodistribution and pharmacological effect of the drug; however the present results indicate that this is not the case. Pharmacokinetics parameters - The results are displayed in Table IV. BNZ presented higher values of Cmax and AUC in blood than MLV-BNZ, whereas both formulations had the same tmax. Since equal doses of BNZ and MLV-BNZ were injected, a similar AUC would be expected. The reason for the lower AUC in blood for MLV-BNZ can be due to the fact that when injected 60% of BNZ remains as liposomal drug. This liposomal BNZ experiences a non-conventional biodistribution, since it is selectively captured by KC in the liver, instead of uniformly diffusing through the vessels to the extravascular space. Another explanation for the lower values of Cmax and AUC in blood exhibited by MLV-BNZ could be the transient increase of vascular permeability produced by local alteration of the veins upon injection by iv of a single bolus of MLV (Rozema & Lewis 2003), reducing the mass of BNZ in the vascular compartment. Another remarkable difference was that Vd was higher for MLV-BNZ than for BNZ, as a direct consequence of its lower association degree to plasma proteins (30% vs 75%), which is inversely related to Vd. The CL corresponding to MLV-BNZ was biphasic, initially fast and ending up at the same rate than that corresponding to free BNZ. The faster rate could be ascribed to the removal of liposomes from blood circulation, exhibiting a t1/2 of 3.2 h, the same order than that required to achieve maximal accumulation in liver upon injection of a single bolus of MLV-BNZ (Morilla et al. 2004). The lower rate corresponds to removal of released BNZ. Hydrophobic drugs such as nystatin, amphotericin B or A cyclosporine extensively associate to plasma proteins; a slight diminution in their association degree substantially increases the amount of drug available to exert pharmacological action (Lin & Liu 1997) and usually alterations in their association degree to lipoproteins also cause modifications in their pharmacological effects (Wasan & Cassidy 1998). However, the hydrophobic BNZ does not extensively associates to plasma proteins; slight changes in its association degree should not substantially increase the percentage of unbound drug available. Additionally, changes in the lipoprotein levels should not be expected to produce alterations in pharmacological activity of BNZ. In relation only to the liposomal formulation, when it was injected by iv, 70% BNZ was available, 60% following a non-conventional and 10% conventional biodistribution, against 25% achieved when the same dose of free drug was injected. These results led to the conclusion that MLV-BNZ is a vehicle potentially capable of improving the pharmacological performance of BNZ and as so should be assayed in animals models of T. cruzi infection. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz. |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}