 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 03, May 2005, pp. 231-235 Triatoma rubrovaria (Blanchard, 1843) (Hemiptera - Reduviidae - Triatominae) IV: bionomic aspects on the vector capacity of nymphs Carlos Eduardo Almeida, Elaine Folly-Ramos*/**, Ronildo Agapito-Souza/*, Gleidson Magno-Esperança/*, Raquel S Pacheco***, Jane Costa Núcleo
de Informatização, Coleção Entomológica,
Departamento de Entomologia ***Departamento de Bioquímica
e Biologia Molecular, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil
4365, 21045-900 Rio de Janeiro, RJ, Brasil *Museu de Ciências,
Centro Universitário de Barrra Mansa, UBM, Barra Mansa, RJ,
Brasil **Programa de Pós-Graduação em Biologia
Animal, Universidade Federal Rural do Rio de Janeiro, Seropédica,
RJ, Brasil +Corresponding author. E-mail: almeidace@ioc.fiocruz.br Received
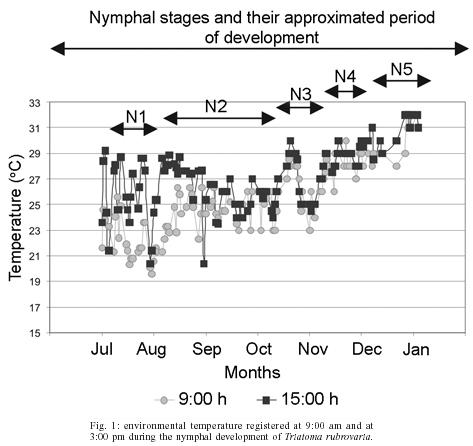
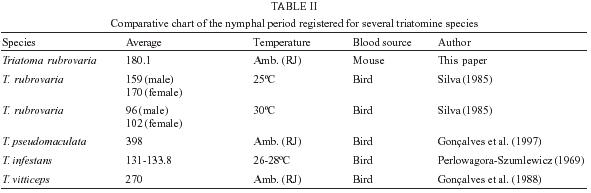
29 November 2004 Code number: oc05074 Triatoma rubrovaria has become the most frequently captured triatomine species since the control of T. infestans in the state of Rio Grande do Sul (RS), Brazil. The aim of this study was to evaluate aspects of the vectorial competence of T. rubrovaria using nymphs raised in laboratory under environmental conditions of temperature and humidity and fed on mice. The average developmental period of T. rubrovaria was 180.1 days. The percentage of defecation shortly after feeding was still higher than previous studies in which samples of T. rubrovaria subjected to a slight starvation period before the blood meal were used. The obtained results support former indication that T. rubrovaria presents bionomic characteristics propitious to be a good vector of Trypanosoma cruzi to man. Therefore its domiciliary invasion process must be continuously monitored. Key words: Triatoma rubrovaria - bionomic studies - vectorial capacity Widespread in Uruguay and parts of Northeastern Argentina, Triatoma rubrovaria (Blanchard, 1843) has its presence confirmed in Brazil only in the southern states of Paraná (PA) and Rio Grande do Sul (RS). This species is mainly found among rocks known as `pedregales' feeding on a wide variety of hosts, including man in perido-miciliary ecotopes (Salvatella et al. 1994, 1995). Utilizing the precipitin test the anti-rodent serum showed the highest positivity from the total of 85 samples collected in several localities. However it is important to mention that 1.28% positivity for human blood was registered (Almeida et al. 2002a). A twenty-year surveillance data gathered by the Brazilian National Health Foundation (Funasa) during the Chagas Disease Control Program show that the presence of T. rubrovaria in human dwellings has been increasing in Southern Brazil (Almeida et al. 2000). Isoenzymatical studies characterized two distinct phenotypic patterns for this species and ecological ones performed in several localities of the RS have also showed food habits differentiation (Almeida et al. 2002a,b). Dias (1956) originally demonstrated that the transmission of Chagas disease depends on the habit of the infected triatomine by Trypanosoma cruzi, in defecating during or shortly after the blood meal. In this context, Almeida et al. (2003) stressed that T. rubrovaria reaches similar index to the ones observed for other effective vector species, such as T. infestans. In the present study other important bionomic information on T. rubrovaria, such as the intermoulting period, the spontaneous fasting period before and after moulting (or hatching), the total number of feedings, and the defecation patterns in subsequent feedings are reported and correlated to its vectorial capacity. MATERIALS AND METHODS Insects
- Samples were taken from a one-year colony kept under laboratory
conditions: temperature (min. 21.5ºC, max. 32ºC,
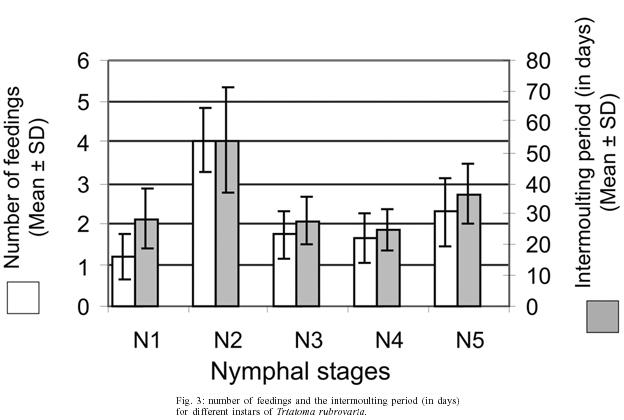
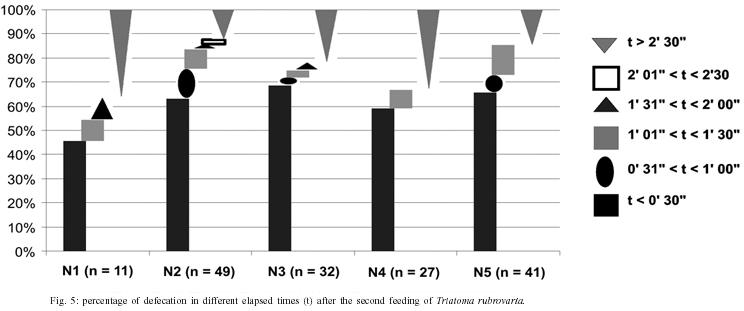
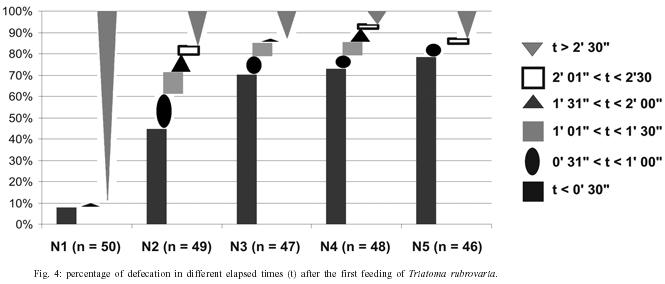
Fifty first nymphs hatched on the same date were randomly picked up from the above mentioned colony. The insects were then promptly transferred to plastic containers (9 × 4 × 4 cm) with perforated lid and containing folded filter paper inside (one insect per container), where they were raised until the adulthood. In this study nymphs from the first to the fifth instar are referred to as N (1 to 5, respectively). According to the method described above, data were collected on: 1 - spontaneous post-ecdysis (or hatching) starvation period [(s)SP-ps] and spontaneous pre-ecdysis starvation period [(s)SP-pr] - for this observation, mice was offered weekly during a period of 10 min. Those nymphs that started feeding were allowed to engorge. Those ones that did not start feeding within 10 min were left for the next meal opportunity until the following week. The number of days that the insect refused a blood meal offer before and after moulting was counted; 2 - number of feedings (NF) - the rate for the mean number of accepted blood meals was calculated individually for each nymphal stage; 3 - intermoult period (IP) - the amount of days between ecdysis for every nymphal stage (N1 to N5) to imaginal emergence was counted in days; and 4 - elapsed time between the end of feeding and defecation - once a week insects were individually transferred to circular plastic arenas (10 cm of diameter on the basis × 15 cm height, with a piece of filter paper as substrate) together with a ketamin anaesthetized mouse (0.35 mg/kg of body weight dose). The elapsed time between the end of feeding and defecation was registered. The end of the blood meal was counted when the proboscis was retracted. When insects defecated during the act of feeding, the time for defecation was set to 0 (immediate defecation). The elapsed time between feeding and defecation at the first (DT1), second (DT2), and third (DT3) feedings of different nymphal stages of T. rubrovaria were also recorded. RESULTS Intermoulting period and bionomic observations - Of the 50 nymphs used at the beginning of the research, only four died before reaching the adulthood: one N2, one N3, and two N4. The
shortest developmental period was in 146 days and longest was recorded
in 226 days ( The lowest average for the (s)SP-ps (2.8 days) was observed in the N1 that also showed the lowest average of NF (1.2). The highest average of (s)SP-ps was registered in the N3 (6.1 days). The N2 showed the highest average of NF (4) and IP (53.8 days). The largest (s)SP-pr was of 21.1 days for N5 and the lowest was of 15.2 days for N4 (Figs 2, 3). In average,
the largest NF in the same stage was observed for the N2 ( Elapsed time between the end of feeding and defecation - The highest percentage of defecation in less than 31 seconds was observed for the N5 (78.2%) at the first feeding. The lowest one was obtained for the N1 (8%), showing an increasing rate from the N1 to N5. With the exception of the N1, we have found that there appears to be a tendency for the maintenance of elevated defecation percentages in less than 31" in all stages after the first feeding, detected on the second and third feedings (Figs 5, 6). The N4 presented the lowest DT1average (41"). Concerning DT2 the lowest average (41") was recorded in the N2. The lowest DT3 (12") was seen in the N4, however these data was obtained from a single sample. This result was followed by the N2 with 01' 29", all these observations were obtained through the analysis of 42 defecations. After DT3 [DT3 to DT(n > 3)], but excluding the N2, just a few specimens defecated in less that 10 min. Thus, the low sampling obtained made it difficult to infer about the averages of elapsed time between the feeding and defecation from the third feeding on (Table III). DISCUSSION Intermoulting period and bionomic observations - Silva (1985) conducted studies on the intermoulting period of T. rubrovaria, by offering bird blood (Gallus gallus) as food source at temperatures of 25ºC and 30ºC. In our study, Mus musculus was utilized as the food source, and the insects were kept under environmental conditions of Rio de Janeiro (Fig. 1). Under these conditions, the average developmental period was 180.1 days. Our result was closed to that reported by Silva (1985) at 25oC (159-170 days). In a former study, Silva (1985) observed that the metabolic condition of T. rubrovaria is strongly influenced by the temperature. A significant statistical difference in the averages of the intermoulting periods at different temperatures was observed: at 25oC the intermoulting period was twice faster than that observed at 30ºC for N1. In our study the average intermoulting period of the N2 was significantly longer than those of any other stages. Under standardized temperature conditions, triatomines trend to exhibit an increasing scale of intermoulting periods from the N1 to N5, as was observed by Silva (1985) for T. rubro-varia. Probably, the variation observed in our study might be due to temperature variations to which the N2 were subjected. During the N2 development a high amplitude of the environmental conditions was observed, while during the summer time the temperature recorded was more stable (Fig. 1). Elapsed time between the end of feeding and defecation - Dias (1956) mentioned that most of the South American analyzed species (Rhodnius prolixus, R. neglectus, T. infestans, P. megistus, T. sordida) defecated in a shorter time after feeding. However, both Dias (1956) and latter Gonçalves et al. (1988) showed that T. vitticeps requires more time to defecate. This same feature was observed for North American species such as T. protracta Ulher (1894), T. rubida (Ulher, 1894) and Paratriatoma hirsuta Barber, 1938, which did not display a defecation pattern conducive to transmission of Chagas disease to man (Wood 1951). Almeida et al. (2003) showed that following a post feeding elapsed time of more than 1 min, most of T. rubrovaria specimens had moved more than 3 cm away from the biting site. Based on these results we conclude that T. cruzi transmission may be a potential risk when the insect defecates either during feeding or in less than 1 min after feeding. Trumper and Gorla (1991) studied the effects of an induced fasting period on the defecation patterns utilizing T. infestans as a model. After moulting the nymphs were kept under starvation regime for 10-12, 19-22, and 24-98 days. The authors concluded that the longer is the fasting period, the longer is the elapsed time to defecate after feeding. For the first feeding, the comparative analysis between the feeding patterns utilizing samples kept under fasting and those ones not starving also displayed defecation patterns substantially different for T. rubrovaria. Almeida et al. (2003) showed that in a regulated fast (20 days of starvation), in the first feeding in each instar, an index of 53.3% (the highest one) of females defecated in less than 1 min after the end of feeding. In this study, 78.2% of N5 displayed its highest index defecating in less than 30" in the first feeding (Fig. 4), when the spontaneous post ecdysis starvation was in mean 5.7 days. Soares et al. (2000) also obtained high indexes of defecation shortly after feeding (50-75%), for nymphs and adults of T. brasiliensis and T. pseudomaculata, which were not submitted to fasting periods. Trumper and Gorla (1991) emphasized that insects kept under long starvation periods required more time to defecate due to necessity of reestablishment of water deficit. These authors state that the fasting period could have a significant implication on T. cruzi transmission, mentioning that nutritional conditions might be affected by the infestation density and that low triatomine population densities could be common either in recently reinfested houses after chemical control or in recent domiciliary colonization. Thus, low population density might also represent high risk for disease transmission. Probably, for T. rubrovaria, if kept under favorable nutritional conditions in the natural environment the values of (s)SP-ps would be similar to those obtained using the methodology presented at our assay. Eliminating the low sampling cases, a general analysis from DT1 to DT(n > 1) indicated that the time elapsed for defecation after the first and the following feedings showed an increasing trend (Figs 4, 5, 6 and Table III) from the fist feeding on. In this work, low mortality in the nymphal period (4 out of 50) was evidenced, suggesting an adaptability of T. rubrovaria to the laboratory conditions. Gonçalves et al. (1988) conducted a similar study with T. vitticeps, and out of the 149 samples used for the biological cycle assay, only 50 reached the adulthood in a mean period of 270 days. All instars of T. rubrovaria showed a number of blood meal in average, higher than that noted for T. vitticeps, T. brasiliensis and R. pictipes, but lower than T. pseudo-maculata (Tables I, II). The obtained data support earlier assumption that T. rubrovaria presents bionomic characteristics that may be favorable to acquire infection and transmit the T. cruzi to man (Almeida et al. 2003). So, the domiciliary invasion process registered for T. rubrovaria must be continually monitored. ACKNOWLEDGEMENT To Dr Terry V Mc Intyre (Centro Unversitário de Barra Mansa) and Dr Pradeep Ghosh, National Institute of Health, for critically reading the manuscript. REFERENCES
The following images related to this document are available:Photo images[oc05074f5.jpg] [oc05074t2.jpg] [oc05074t3.jpg] [oc05074f6.jpg] [oc05074f2.jpg] [oc05074f1.jpg] [oc05074f3.jpg] [oc05074f4.jpg] [oc05074t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}