 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
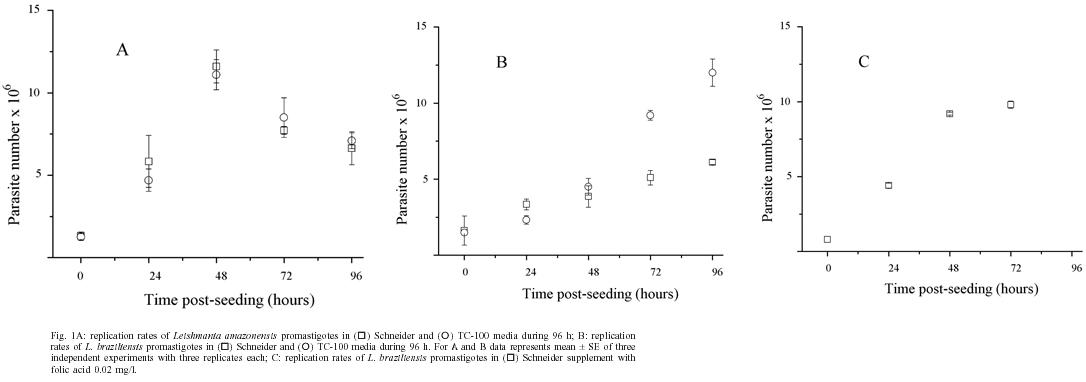
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 3, May 2005, pp. 309-310 Leishmania (Viannia) braziliensis growth
in vitro culture relies more on folic acid availability than Leihsmania Andrea Niño, Marcela Camacho*/+ Laboratorio de
Biofísica, Centro Internacional de Física, Bogotá, Colombia
*Departamento de Biología, Facultad de Ciencias, Universidad Nacional
de Colombia, Ciudad Universitaria, Calle 45, Carrera 30 Bogotá, Colombia Programa Nacional
de Ciencia y Tecnología de la Salud, División de Investigaciones
project 90835, Universidad Nacional de Colombia, Centro Internacional de
Física Received 13 October
2004 Code number: oc05089 We compared the in vitro growth of promastigotes from two Leishmania species in TC-100 and Schneider media. Leishmania (Leishmania) amazonensis replication rates were similar in both tissue culture media and reached maximum rates by 48 h. In contrast Leishmania (Viannia) braziliensis growth was significantly greater in TC-100 but maximum rates were achieved by 96 h. Folic acid appears to be the limiting factor and supplementation of Schneider media with this nutrient improved L. (V.) braziliensis replication rates and decreased the time of maximum replication to 48 h. Key words: Leishmania (Viannia) braziliensis - Leishmania (Leishmania) amazonensis - replication rates - in vitro culture. Leishmania are parasitic protozoa that infect many mammals including man. Despite the epidemiological impact of Leishmania (Viannia) braziliensis in the New World (OPS 1996), it has been studied less than other species, principally due to the difficulties of keeping this parasite in axenic conditions. These difficulties were used in the past to differentiate this Leishmania species and were related to media composition differences (Walton et al. 1977, Shaw & Laison 1981). Biphasic media with solid phases enriched with variety of components have been used to keep Leishmania, but harvesting large numbers of parasites is difficult, thus liquid media is preferred for promastigote cultivation (Schuster & Sullivan 2002). L. (V.) braziliensis promastigotes show low growth rates in in vitro cell culture, when they are compared with other Leishmania genus species (Armstrong & Paterson 1994). In this study we compare the ability of two different media to sustain L. (V.) braziliensis and L. (L.) amazonensis growth. MATERIALS AND METHODS Parasites were cultured as previously described (Forero et al. 1999). L. (V.) braziliensis (HOM/BR/75M2903) and L. (L.) amazonensis (FLA/BR/67/PH8) isolates, kindly donated by Dr Nancy Gore Saravia (Cideim, Cali, Colombia) were used in the present study. Promastigotes, at an initial concentration of 1 x 106, were cultured at 24oC in 25 cm2 flasks (Costar) in TC-100 or Schneider media (In-vitrogen) supplemented with 10% FBS (Hyclone). Cultures were allowed to reach their metacyclic phase, determined by the stationary behavior of parasite number and rosette formation. Samples of parasites at different time points were counted in a Neubauer hematocytometer in a solution of 5% Giemsa, 2% formaldehyde in 0.14 M NaCl, under light microscopy. Data were analyzed with the Student's t test and p values < 0.05 were considered significant. RESULTS L. (L.) amazonensis replication rates were similar in the two culture media during the period studied, and maximum replication rates occurred within 48 h (Fig. 1A). L. (V.) braziliensis replication rates doubled during the first 48 h and no differences were found between the two culture media (p = 0.4). However, between 72 to 96 h post-seeding, the replication rates of L. (V.) braziliensis promastigotes tripled in TC-100 but only doubled in Schneider. This difference was significant (p = 0.02; Fig. 1B). Although L. (V.) braziliensis took 96 h to reach its optimal replication rate compared to L. (L.) amazonensis, parasite numbers were similar. The percentage of metacyclics increased over the period studied for both species, and achieved their maximum value during the stationary phase. To further investigate the impact of particular nutritional factors on the recorded differences, folic acid was added to Schneider media. L. (V.) braziliensis replication rates increased and the maximum replication time was reduced to 48 h post-seeding, suggesting that the limiting nutritional factor was folic acid (Fig. 1C). DISCUSSION We found no differences between replication rates of L. (L.) amazonensis promastigotes in the two media studied. In contrast L. (V.) braziliensis promastigote replication rates were significantly higher in TC-100 compared to Schneider media. This difference may be the result of the presence of a toxic component in Schneider that is absent in TC-100 media, that L. (V.) braziliensis is more susceptible to. On the other hand, it could be the result of the depletion of an essential component in Schneider media that L. (V.) braziliensis needs. We rule out the first explanation because no differences were observed in the ability of these two media to sustain similar replication rates of this parasite during the first 48 h of culture. Nutrient requirements in Leishmania indicate that these parasites are folate and pterin auxotroph, but the requirements for these nutrients varies among Leishmania species. In L. (V.) braziliensis, this is particularly important since it has been reported that higher concentrations of folic acid are required by this species compared to other Leishmania (Steiger & Steiger 1977). Folic acid has been implicated in tymidine and methionine synthesis and in the interconversion of serine into glycine by Leishmania (Ouellete et al. 2002). It was also showed that absence of folic acid inhibited L. (L.) donovani growth (Schuster & Sullivan 2002). Therefore, we favor the depletion of an essential component as a more likely explanation. Schneider contains important amounts (2 g/l) of a non defined component yeastolate, but lacks folic acid as a defined component. On the other hand, TC-100 contains folic acid at micromolar concentrations (Gardiner & Stockdale 1975), in the order or higher than those required by L. (L.) donovani (Peixoto & Beverley 1987) and L. (L.) major (Ellenberger & Beverley 1987). Also, the fact that L. (V.) braziliensis replication rates improved and parasite growth was faster after the addition of folic acid to Schneider media adds further support to the hypothesis that depletion was responsible for the differences between the two media after 48 h. REFERENCES
The following images related to this document are available:Photo images[oc05089f1.jpg] |
| |||||||||

{kind=link}