 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 3, May 2005, pp. 319-324 Slime production and proteinase activity of Candida species isolated from blood samples and the comparison of these activities with minimum inhibitory concentration values of antifungal agents Semiha Ozkan/+, Fatma Kaynak, Ayse Kalkanci*, Ufuk Abbasoglu, Semra Kustimur* Department of Pharmaceutical
Microbiology, Faculty of Pharmacy *Department of Microbiology, Faculty of
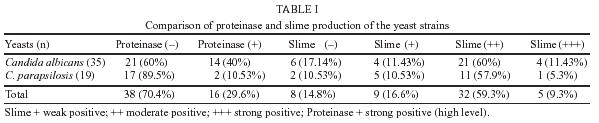
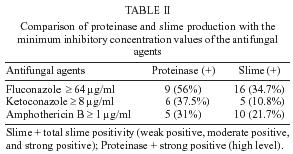
Medicine, Received 29 October 2004 Code Number: oc05091 Slime and proteinase activity of 54 strains consisting of 19 Candida parapsilosis and 35 C. albicans strains isolated from blood samples were investigated in this study. Ketoconazole, amphothericin B, and fluconazole susceptibility of Candida species were compared with slime production and proteinase activity of these species. For both Candida species, no correlation was detected between the slime activity and minimum inhibitory concentration (MIC) values of the three antifungal agents. For both Candida species no correlation was detected between the proteinase activity and the MIC values of amphothericin B, and fluconazole however, statistically significant difference, was determined between the proteinase activity and MIC values of ketoconazole (p = 0.007). Slime production was determined by using modified Christensen macrotube method and proteinase activity was measured by the method of Staib. Antifungal susceptibility was determined through the guidelines of National Committee for Laboratory Standards (NCCLS M27-A). Key words: slime
- proteinase activity - Candida spp. - antifungal susceptibility -
minimum inhibitory concentration - During past two decades, Candida infections have increased in number and severity (Zaugg et al. 2001). The increase in Candida has been accompanied by a significant rise in morbidity and mortality (Pichová et al. 1999) and Candida blood stream infection have been associated with these highest rates of morbidity and mortality (Edmond et al. 1999). In immunocompromised patients, Candida species have the potential to invade all host organs and cause severe systemic infections. Although C. albicans is the organism most often associated with serious fungal infections, other Candida species have emerged as clinically important pathogens of these opportunistic infections. C. parapsilosis is the second most important pathogenic yeast of the Candida species (Kuhn et al. 2002b). Candida strains possess a number of virulence factors those enable the organism to cause hematogenously disseminated infections in susceptible hosts which aid persistence and colonization of the host tissue. These include the slime production, to undergo a dimorphic transition and to produce the extracellular proteinase (Ibrahim et al. 1993, Kretschmar et al. 1999). Biofilms represent the most prevalent type of microbial growth in nature and are crucial to the development of clinical infections. In the latter setting, they are associated with high-level antibiotic resistance of the associated organisms. Candida is the fourth most common cause of bloodstream infections in hospitalized patients. In several studies, C. parapsilosis has become the second most commonly isolated fungal organism (Kuhn et al. 2002a). In this study the slime and proteinase activity of Candida species isolated from blood samples were determined and the results were compared with MIC values of ketoconazole, amphothericin B, and fluconazole. MATERIALS AND METHODS Yeasts - Thirty five C. albicans and 19 C. parapsilosis strains were isolated from blood samples. The isolates were provided from Gazi University Faculty of Medicine Microbiology Laboratory. All the yeasts were isolated from different patients. Blood cultures were performed using Bactec NR 660 and aerobic plus bottles (10 ml of blood). The yeast isolates were identified in terms of their gross morphological, microscopic, and physiologic characteristics according to the methods recommended by Kurtzman and Fell (1998). C. albicans ATCC 10231 reference strain was also included in this study. Media - Sabouraud liquid medium (Merck) containing 4% glucose, Bactoagar (BA) (Merck), Sabouraud liquid medium (SLM) (Merck), Agar agar (Oxoid) were used in this study. Chemical agents - 0.1%[w/v] KH2PO4 (Merck), 0.5%[w/v] MgSO4(Merck), 1%[w/v] glucose, 0.16%[w/v] bovine serum albumin (Sigma), 20% trichloroacetic acid, 1.25% amidoblack (90% methanol, 10% acetic acid), Dimethylsulfoxide (DMSO) (Merck), Phosphate buffer solution (PBS) (pH=7.2) were used during the examination. Antifungal agents - The susceptibility of fluconazole (Fako, Turkey), ketoconazole (Bilim, Turkey) and amphothericin B (Bristol, Myers Squibb, UK) were determined to compare with the slime and proteinase activities. Antifungal susceptibility testing - The MIC of antifungal agents for all isolates were determined by broth microdilution assays for yeasts according to the National Committee for Clinical Laboratory Standards (NCCLS-M27A) guidelines for yeasts (NCCLS 1997). Slime production - Slime production was determined by using a modification of the test established for coagulase-negative staphylococci (Christensen et al. 1982). A loopful of organisms from the surface of a Sabouraud Dextrose agar plate was inoculated into a tube containing 10 ml Sabouraud liquid medium supplemented with glucose (8%). The tubes were incubated at 35°C for 24 h after which the broth was aspirated and the walls of the tube were stained with safranin. The adherent slime layer was scored as negative, weak positive (1+), moderate positive (2+ or 3+) or strong positive (4+). Each tube was scored independently by two observers. Proteinase assay - Proteinase production was measured by the method of Staib (1965). Cultures were grown overnight in SLM at 36°C and 200 rpm in an orbital incubator. Cells were harvested by centrifugation, washed with PBS and resuspended at a density of 1x108 cells/ml. 10 µl amounts were placed onto the test medium [1% agar, 0.1%[w/v] KH2PO4, 0.5%[w/v] MgSO4, 1%[w/v] glucose] containing 0.16%[w/v] bovine serum albumin as the sole nitrogen source. After incubation for five days at 37°C, the plates were fixed with 20% trichloroacetic acid and stained with 1.25% amidoblack. Decolorization was performed with 15% acetic acid and the clear zones around each colony were measured and used in the determination of the precipitation zone (Pz) values. Assays were performed in replicates of three, on two separate occasions. The Pz indexes were determined as described by Price et al. (1982). Pz = 1-A/B where "A" is the diameter of the colony + zone of precipitation and "B" is the diameter of the colony. When Pz was 1.00, enzymatic activity was zero. Between 0.64 and 0.99, this activity was considered positive, when lower than 0.64 it confirmed a strong positivity. Statistical methods - Proteinase and slime activities were compared by the Mann-Whitney test for independent samples. Chi-square analysis was used to test the correlation of proteinase and slime activity groups with antifungal susceptibility of antifungal agents. p < 0.05 was considered significant. RESULTS Slime and proteinase activity of 54 strains consisting of 19 C. parapsilosis and 35 C. albicans strains isolated from blood samples were investigated in this study. Of the 54 strains, slime production was found in 46 strains tested (80.5%), 82.8% of C. albicans strains were slime positive while 89.4% of C. parapsilosis strains showed this activity. Weak slime production was found in 9 strains (19.5%) and strong slime production was found in 5 strains (11%). Proteinase activity was found in 40% of C. albicans strains and 10.5% of C. parapsilosis strains showed the same activity. There was no statistically significant difference between C. albicans and C. parapsilosis in terms of slime production (p = 0.4) however the difference was significant in terms of proteinase activity (p = 0.015). The results are shown in Table I. We investigated the correlation between slime and proteinase activity with MIC values of ketoconazole, amphothericin B, and fluconazole. Of the 46 slime positive strains, 34.7% were found as resistant to fluconazole, 21.7% amphothericin and 10.8% ketoconazole.The values of MIC were ³ 64 µg/ml, ³ 1 µg/ml, and ³ 8 µg/ml for fluconazole, amphothericin, and ketoconazole, respectively. For both Candida species no correlation was detected between the slime activity and MIC values of the three antifungal agents. Proteinase activity was found in 16 of the total of 54 strains (29.6%) and the activity was calculated as high level (Pz < 0.64) in all positive strains. Of the 16 proteinase positive strains, 56% were found as resistant to fluconazole (MIC > 64 µg/ml) and MIC values of 31% were ³ 1 µg/ml for amphothericin B and 37.5% were ³ 8 µg/ml for ketoconazole. For both Candida species no correlation was detected between the proteinase activity and the MIC values of amphothericin B and fluconazole but statistically significant difference was observed between the proteinase activity and ketoconazole (Table II). As a result, while the slime activity was present for more strains than proteinase activity, the correlation between the MIC values and proteinase activity was more significant. DISCUSSION Since 1990, it has become clear that yeasts of the genus Candida continue to be important etiologic agents of nosocomial blood infections. Furthermore, the proportion of such infections due to C. non-albicans species is persistently rising (Fridkin & Javis 1996). Such increase was particularly observed in a pioneering study of Farina et al. (1999) carried out during the period of 1988 through 1997 at the Regional Hospital of Bergamo, in Italy. The authors analyzed 168 cases of fungemia, of which 70% were due to Candida spp. Of these, 41.4% were identified as C. albicans and 37.9% as C. non-albicans. This shows the need for a correct identification of Candida isolates at the species level (Pfaller 1998). Additionally, there must be a continuous surveillance of the standards of antifungal susceptibility, as they are important components of any program intended to prevent and control nosocomial fungal infections (Pfaller et al. 1997). A possible explanation for the emergence of C. non-albicans is the selection of more resistant species through the use of antifungal agents (Pfaller et al. 1995a). Virulence factors of Candida spp. have been of great interest as targets for the development of new therapeutic interventions against candidiasis (Perfect et al. 1996). Such factors include mainly, proteinases (Hube et al. 1998) and biofilms (Rawi & Kavanagh 1999). Extracellular proteases of eucaryotic microbial pathogens have attracted the attention of many laboratories because of their potential role in pathogenesis. They facilitate the penetration into the host organism and counteract its defence system. Analysis of proteolytic enzymes of pathogenic microorganisms might lead to a design of inhibitors controlling these pathogens. A considerable amount of information is now available on the extracellular proteases of fungi, especially of the genera Candida. C. albicans, the major human pathogen of the genus Candida, is a commensal organism in healthy individuals and seems to be almost universally present. Superficial infections, especially those of the mouth and vagina, are very common and often prove refractory to a treatment. Among the virulence factors of C. albicans, the production of acid proteinases has been extensively studied in view of its important role in the pathogenesis of candidiasis. The role of secreted aspartic proteases of C. albicans in experimental and clinical candidiasis has been demonstrated. The enzymatic production of C. albicans and other species isolated from different clinical conditions and anatomical sites was studied and variations of 62.5-100% for proteinase activity were determined (Ruchel et al. 1982, 1983, Samaranayake et al. 1984, Maffei et al. 1997, Pichova et al. 2001). However, other species of the genus Candida, such as C. tropicalis, C. parapsilosis, C. glabrata, C. krusei, and C. lusitaniae, have been shown to cause an increasing incidence of mycoses. These yeasts are often associated with resistance to antifungal azoles and with higher mortality (Pichova et al. 2001). Bernardis et al. (1999) found that cutaneous isolates of C.parapsilosis had uniformly elevated secretory aspartyl-protease activity, more than four times higher than the enzyme activity of the blood isolates. To avoid the effect isolate source, we used only blood isolates in this study. This result suggests that extracellular proteinase activity is correlated with another factor, such as source of the isolate. In our study, C. albicans had significantly higher proteinase activities (40%), than C. parapsilosis (10.53%) (p < 0.015). Matsumoto et al. (2001) isolated 80 yeast strains from hospitalized children, being 59 from blood cultures and 21 from vascular catheter cultures. The prevalent species in both blood and catheter was C. parapsilosis (32.2 and 48.9%, respectively), followed by C. albicans (16.9 and 28.6%, respectively). Concerning enzyme production, 78.8% of the 80 isolates presented strong proteolytic activity. The mean secretory proteinase activity of C. albicans isolates from the oropharyngeal candidiasis in HIV positive group was significantly higher when compared with the isolates from the control group (p < 0.05). Proteinase activity within the population of C. albicans isolates from HIV positive individuals was independent of the patient's clinical disease stage and no correlation of the proteinase activities with the C. albicans serotype was found. However a positive correlation of proteinase activity to antifungal susceptibility was evident. The C. albicans isolates from the HIV positive group that were characterized by higher levels of proteinase activity were also less susceptible to the widely used azole antifungal agents ketoconazole and fluconazole (Ollert et al. 1995). A biofilm is a community of microorganisms and their extracellular polymers that are attached to a surface (Pfaller et al. 1995). Biofilms represent the most prevalent type of microbial growth in nature and are crucial to the development of clinical infections (Branchini et al. 1994). While previous studies of biofilm development and species interaction have focused largely on bacterial species, relatively little is known about fungal biofilms. C. albicans biofilms share several properties with bacterial biofilms, including their structural heterogeneity, the presence of expolymeric material, and their decreased susceptibility to antimicrobial agents and biocides (Ramage et al. 2001a). The ability to form extensive biofilms on the surface of catheters, and other prosthetic devices, also contributes to the high prevalence of the organism as etiologic agent of intravascular nosocomial infections (Matsumoto et al. 2001). Biofilm production was also associated with high-level antimicrobial resistance of the associated organisms. Candida is the fourth most common cause of bloodstream infections in hospitalized patients and in several studies C. parapsilosis has become the second most commonly isolated fungal organism. Slime production may be important in enabling C. parapsilosis to cause catheter-related bloodstream infections because C. parapsilosis is an important nosocomial pathogen that can proliferate in high concentrations of glucose and form biofilms on prosthetic materials (Branchini et al. 1994). Kalkanci et al. (1999) investigated the correlation between the Candida strains and clinical isolation areas for slime production. Weak slime production was found in 8% and strong slime production was found in 4% of the strains. Slime positivity for C. albicans was 9.3%; on the other hand it was 25% for non-albicans strains. The slime factor is more important for non-albicans strains (C. kefyr, C. tropicalis, C. rugosa, C. krusei). In this present study, there was no statistically significant difference between C. albicans and C. para-psilosis in terms of slime production (p = 0.4). Biofilms, likely the predominant mode of device-related microbial infection, exhibit resistance to antimicrobial agents. Evidence suggests that biofilms have dramatically reduced susceptibility to antifungal drugs (Kuhn et al. 2002b). Consequently, biofilm-related infections are inherently difficult to treat and to fully eradicate with normal treatment regimens. Antifungal susceptibility testing represents a means of predicting therapeutic concentrations of antifungal drugs used to treat a variety of Candida infections (Ramage et al. 2001b). Pfaller (1995) investigated the slime production and antifungal susceptibility among 60 isolates of C. parapsilosis. Antifungal susceptibility to amphothericin B, 5-fluorocytosine, fluconazole, and itraconazole was determined. It was observed that the association between slime production and resistance. Yücesoy et al. (2003) have investigated the biofilm production of various Candida strains and compared this activity with fluconazole and amphothericin B susceptibility. There was no statistically significant difference between Candida strains biofilm activity and susceptibilities to amphothericin B (p = 0.12), however, with Candida strains statistically significant difference was determined between biofilm activity and susceptibilities to fluconazole by tube adherence method (p = 0.03). In our study, 19 C. parapsilosis and 35 C. albicans strains isolated from blood samples were investigated whether the proteinase and slime activity patterns correlated with the strains' susceptibilities to ketoconazole, fluconazole and amphothericine B. For both Candida species, no correlation was detected between the slime activity and MIC values of all three antifungal agents. However for both Candida species statistically significant difference was determined between the proteinase activity and MIC values of ketoconazole (p = 0.007) (Table II). C. albicans isolates (65.7%) were resistant ( ³ 64 µg/µl) to fluconazole and among them 56.25% showed acid proteinase production and 60% showed strong positive slime activity. 22.3% of C. albicans isolates were resistant (³ 8 µg/ml) to ketoconazole and of these isolates 37.5 % showed acid proteinase production and 40% showed strong positive slime activity. Fluconazole and ketoconazole resistance of ³ 8 µg/ml in C. parapsilosis isolates was not detected and for three C. parapsilosis isolates the susceptibility was determined as ³ 1 µg/ml for amphothericin B. As our results, while the slime activity was determined for more strains than proteinase activity, the correlation between the MIC values and proteinase activity was more significant. It has been thought that the proteinase activity increases resistance to azole group antifungal agents especially in immunosuppressed patients. These results demonstrated that more studies are needed to show the proteinase activity and antifungal susceptibility testing in this group of patients. REFERENCES
Copyright 2005 Instituto Oswaldo Cruz - Fiocruz The following images related to this document are available:Photo images[oc05091t3.jpg] [oc05091t1.jpg] [oc05091t2.jpg] |
| |||||||||

{kind=link}
{kind=link}