 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 3, May 2005, pp. 325-330 Benznidazole-induced genotoxicity in diploid cells of Aspergillus nidulans Edilson N. Kaneshima, Marialba A. Alves de Castro-Prado*/+ Universidade Estadual
de Maringá, Departamento de Análises Clínicas *Departamento
de Biologia Celular e Genética, Av. Colombo 5790, 87020-900 Maringá,
PR, Brasil Received 18 December
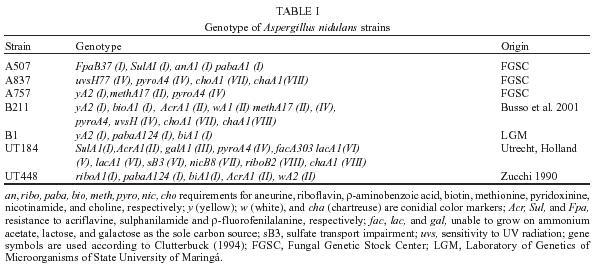
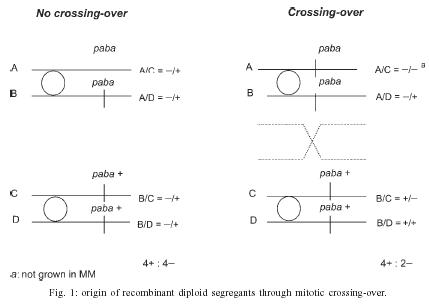
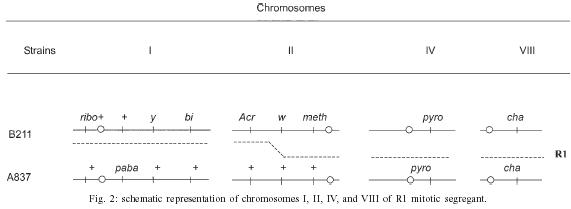
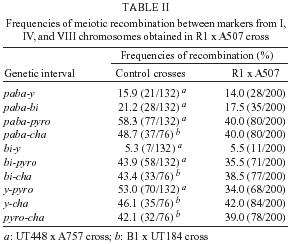
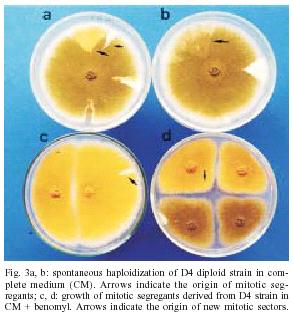
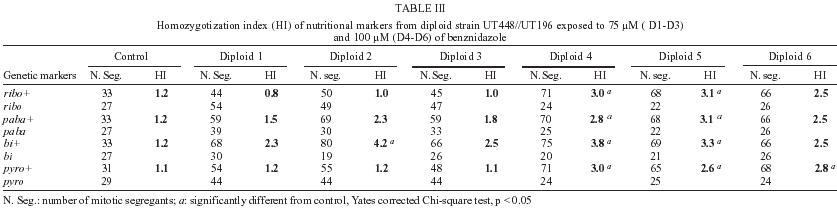
2004 Code number: oc05092 Genotoxic effects of benznidazole were studied by the induction of homozygosis of genes previously present in heterozygous. UT448//A757 diploid strain was used in the benznidazole's recombinagenesis test. Although toxic effects on growth of colonies were not observed, 75 and 100 µM benznidazole induced an increasing of mitotic recombination events in diploid strain. Results were related to the induction of chromosomal breaks by the antiparasitic drug. Key words: benznidazole - somatic recombination - genotoxicity - homozigotization index Chagas disease, caused by Trypanosoma cruzi, is one of the most serious parasitic diseases of Latin America, where some 16 - 18 million people are infected by the T. cruzi (WHO 1991). Although in Brazil the disease is under strict control, there are still about 4 million people infected by the parasite (Dias 1998). Benznidazole is used in the treatment of Chagas disease in the acute phase, at the start of the chronic phase, in congenital cases and accidental infection (Fragata Filho et al. 1997). Several adverse effects have been described in patients undergoing treatment with the anti-parasitic drug (Cançado 1997, Fragata Filho et al. 1997). The mutagenic action of the benznidazol has been studied in streptomycin-resistant Klebsiella pneumoniae strain and in Salmonella typhimurium TA100 strain. Mutagenic effect of the drug has also been observed in TA100 strain in the presence and in the absence of the microsomal hepatic fraction of mice, using 50 to 1000 µM concentrations (Voogd et al. 1975, Nagel & Nepomnaschy 1983). The mutagenic activity of benznidazole has also been determined in the simplified version of Ames test with sensitive (TA100 and TA102) and less sensitive (TA98 and TA97) S. typhimurium strains (Melo & Ferreira 1990) in the same plate. The drug's mutagenic effect was accounted to the interaction of reactive metabolites of benznidazole with the DNA and with nuclear proteins (Gorla et al. 1986, Zahoor et al. 1987, Ferreira et al. 1988). Benznidazole's genotoxicity has been studied in peritoneal macrophages of mice treated with benznidazole (Lacava & Luna 1994). These authors reported the clastogenic effect of the drug owing to increase in the frequency of chromatid breaks and to the formation of acrocentric chromosomes. Moya and Trombotto (1988) demonstrated the ben-znidazole`s clastogenic effect in patients treated with the drug. High micronucleus percentage and increase in the frequency of fragile sites (gaps or breaks in the metaphase chromosomes) were observed in cytogenetic analysis. Bocchi et al. (1998) detected a high incidence of malignant neoplasms in patient with Chagas disease who had undergone heart transplant and treated with benznidazole. On the other hand, although Teixeira et al. (1994) observed a high incidence of lymphoblastic lymphoma in mice treated with benznidazole, Andrade et al. (2003) verified that the administration of the benznidazole in immunosuppressed mice and chronically infected by T. cruzi did not result in the emergence of lymphomas or other neoplasms. In spite of these contradictory results, the clastogenic effect of benznidazole (Lacava & Luna 1994) suggest its participation in the carcinogenic process due to the occurrence of mitotic crossing-over that induces the loss of heterozygosity of tumor suppressor genes (Zimmermann 1971, Weinberg 1991, Beumer et al. 1998). The evaluation of the recombinagenic potential of benznidazol in heterozygous cells of Aspergillus nidulans will be provided. Information on the participation of the anti-parasitic drug in the carcinogenesis process will be thus ensued. MATERIALS AND METHODS Strains and culture media - A. nidulans strains used are described in Table I. Minimum medium (MM) was Czape-Dox with 1% (w/v) glucose. Complete medium (CM) has previously been described by Pontecorvo et al. (1953) and Van De Vate and Jansen (1978). Supplemented medium (SM) consisted of MM plus nutriments required by each strain. Solid medium contained 1.5% agar. Incubation occurred at 37oC. Methods - General methodology followed previous reports (Roper 1952, Pontecorvo et al. 1953). Heterokaryons were prepared in liquid MM plus 2% CM. Cleistothecia were obtained from heterokaryons after 21 days of incubation in sealed petri dishes containing MM, supplemented according to the requirements of the crossed strains. Diploids were prepared by method described by Roper (1952). Evaluation of drug toxicity - Filter-sterilized aqueous solutions of benznidazole (Roche), 99.8% pure, was added to molten CM so that final 50, 75, and 100 µM concentrations could be obtained. Conidia of diploid strains UT448/A757 and B211//A837 were inoculated in plates containing CM (control) and CM + benznidazole (treatment). Five plates, were used for each dose and for control. Diameters of colonies were measured after 24, 48, 72, 96, and 120 h of incubation. Comparison of values of colony diameters with and without the drug was undertaken by Student's t test. Evaluation of the drug' recombinagenic potential - Conidia of diploid strain UT448//A757 were inoculated in plates with MM + benznidazole (50, 75, and 100 µM). Plates were incubated for 6 days at 37°C. Treatment produced visible diploid sectors, D1-D6, identified by differentiated morphology of the original diploid. Sectors were homozygous (+/+) or heterozygous (+/- or -/+) diploid segregants, but never recessive homozygous (-/-), since they did not grow in MM. Diploid sectors were haploidized spontaneously in CM after purified in MM. After haploidization the haploid mitotic segregants from diploids D1 to D6 were purified in CM. Only mitotic stable segregants were selected for the recombinagenic test and determination of the homozigotization index (HI) (Pires & Zucchi 1994, Franzoni et al. 1997). Determination of the homozigotization index (HI) - Conidia of each haploid segregant were transferred, one-by-one, to 25 positions in CM plates (master plates). After incubation for 48 h at 37ºC, colonies were transferred to the proper selective media for phenotypic analysis of haploid segregants. Nutritional markers of the haploid segregants segregate in the proportion (a) 4+:4-, if drug fails to induce recombinagenesis or (b) 4+: 2-, if drug induces crossing-over (Fig. 1). The homozygotization index (HI), or the ratio between prototrophic and auxotrophic segregants, equal to or over 2.0 (4+/2-), reveals the recombinagenic effect of the compound under analysis (Pires & Zucchi 1994, Chiuchetta & Castro-Prado 2002, Busso & Castro-Prado 2004). RESULTS AND DISCUSSION Three different benznidazole concentrations were assayed for their ability to induce cytotoxic effects and somatic segregation in the A. nidulans mould. Benznidazole had no effect on colonies' morphologies and micelial growth of diploid UT448//A757 and B211//A837 strains at the three tested concentrations (results not shown). Induction of aneuploidy and mitotic crossing-over was studied in heterozygous diploid B211//A837 strain exposed to benznidazole. Treatment of B211//A837 with benznidazole 100 mM in CM allowed the isolation of a mitotic segregant named R1. Phenotypic analyses of R1 showed it was recombinant for Acr-w interval of chromosome II (Fig. 2). Segregant was grown in CM + benomyl (2 µg/ml) so that its mitotic stability could be analyzed. R1 did not produce new mitotic sectors with the haploidization agent and was classified as haploid segregant (results not shown). In fact, when submitted to the sexual cycle, R1 produced normal frequencies of meiotic recombination for markers of chromosomes I, IV, and VIII (Table II). Prototrophic diploid segregants were isolated from UT448//A757 colonies, after treatment with 50, 75, and 100 µM of benznidazole in MM. Diploids (D1 to D9) were submitted to spontaneous haploidization in CM (Fig. 3a,b) and the selected mitotic segregants were tested for their mitotic stability in CM + benomyl (Fig. 3c,d). Only segregants that failed to produce new mitotic sectors, demonstrating genetic stability, were selected for HI determination. Although HI values obtained from benznidazole 50 µM were lower than 2.0 (results not shown), results obtained with 75 and 100 µM demonstrate that benznidazole is effective in inducing mitotic crossing-over in A. nidulans diploid strain. HI values obtained from benznidazole 75 and 100 µM (D2, D4, D5, and D6) were higher than 2.0 and statistically different from control (Table III). The recombinagenic effect of benznidazole may be related to the induction of chromosomal breaks, such as has been observed in cytogenetic analyses of human lymphocytes and of mice's peritoneal macrophages (Moya & Trombotto 1988, Lacava & Luna 1994). Multiple genetic alterations, such as point mutations, chromosomal translocations and loss of heterozygosity (LOH), are involved in the cellular carcinogenesis (Barrett 1993, Ramel et al. 1996). In human retinoblastoma, LOH is the most common mechanism by which the normal wild-type allele at the RB1 locus is lost in a heterozygous retinal cell for a null mutation. Possible chromosomal mechanism triggering LOH would include: mitotic non-disjunction with loss of chromosome bearing the wild-allele or mitotic recombination between the RB1 locus and the centromere, resulting in homozygosity of defective allele (Cavenee et al. 1991, Hagstrom & Dryja 1999). Somatic recombination consists of exchange events between homologous chromosomes that, following chromosome segregation and cell division, may result in homozygosis of distal genes to the point of exchange (Lasko et al. 1991, Zimmermann 1992, Beumer et al. 1998). Although the rate of spontaneous mitotic recombination in dividing cells of mammals is very low (Morley et al. 1990), it is known that mitotic recombination may occur during the repair of chromosomal double-strand and single-strand break (Galli & Schiestl 1998, Hagstrom & Dryja 1999, Helleday 2003, Stark & Jasin 2003). Benznidazole in vitro increased the frequency of the sister chromatid exchanges in human cells of hepatoma and in lymphocytes of patients treated with the drug. Drug is further capable of increasing the micronuclei frequency in the hepatoma cells (Santos et al. 1994). The clastogenic effect of the anti-parasitic agent was also observed in peripheral lymphocytes of chagasic children (Gorla 1988). Although the mutagenic effect of benznidazole at 50 µM has been observed in S. typhimurium assays (Nagel & Nepomnaschy 1983), this dose does not show re-combinagenic effect in A. nidulans (results not shown). Our results demonstrate that benznidazole recom-binagenic effect is dose-dependent. Since somatic recombination may trigger neoplasms, current analysis suggests that the carcinogenic potential of the anti-parasitic drug may be conducted by loss of heterozygosity mediated by mitotic crossing-over. ACKNOWLEDGMENTS To Dr Francisco Gualter for supplying benznidazole (Roche) and to Luzia AS Regasse for technical assistance. REFERENCES
The following images related to this document are available:Photo images[oc05092t2.jpg] [oc05092f3.jpg] [oc05092f1.jpg] [oc05092t3.jpg] [oc05092t1.jpg] [oc05092f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}