 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 3, May 2005, pp. 331-337 Lymphocyte subset analyses in healthy adults vaccinated with yellow fever 17DD virus Ana Paula dos Santos/++, Álvaro Luiz Bertho*, Daniela Capuzzo Dias/++, Jaciara Ramos Santos*, Rugimar Marcovistz/+ Laboratório de Tecnologia Imunológica, Bio-Manguinhos-Fiocruz *Laboratório de Imunoparasitologia, Departamento de Imunologia, IOC-Fiocruz, Av. Brasil 4365, 21045-900 Rio de Janeiro, RJ, Brasil +Corresponding
author. E-mail: rugimar@bio.fiocruz.br Received 28 July
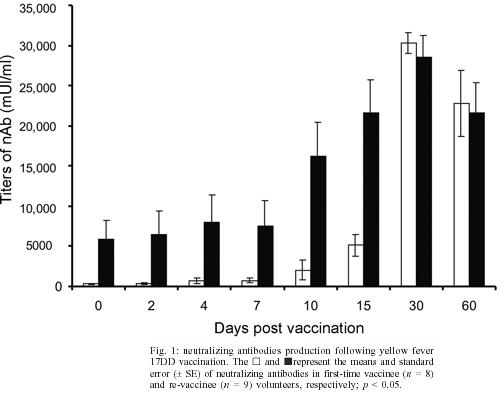
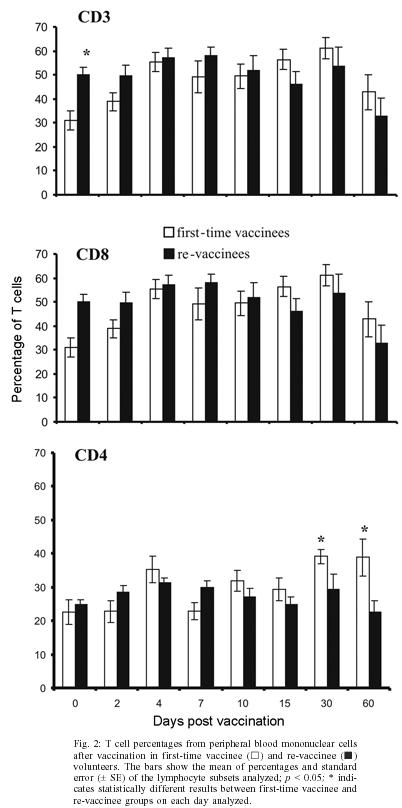
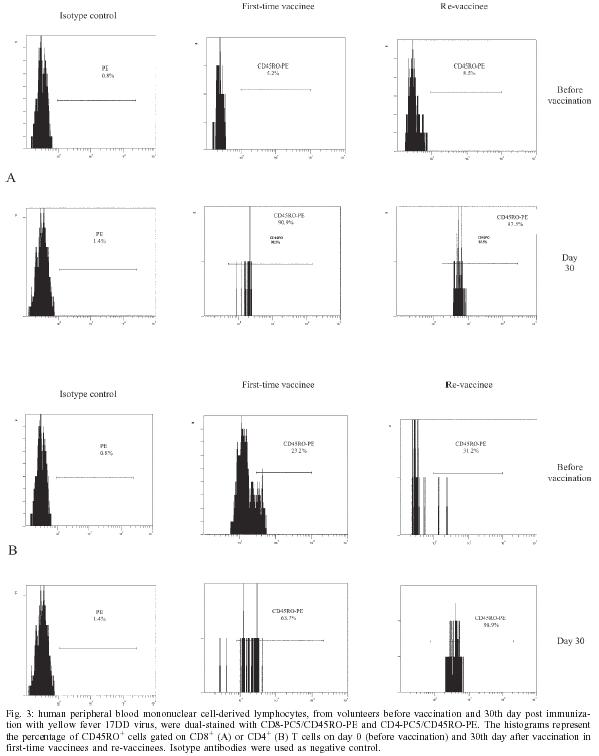
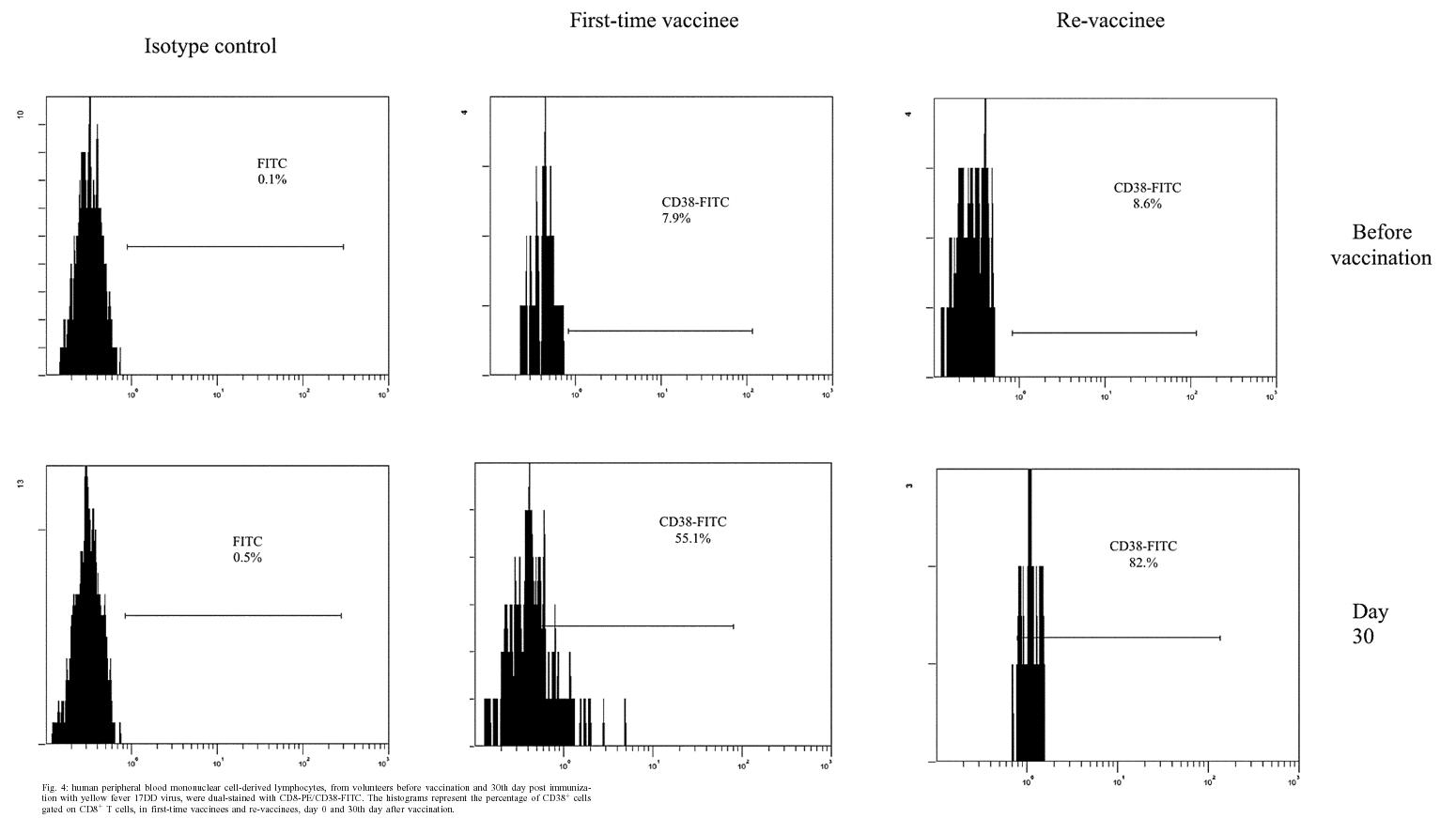
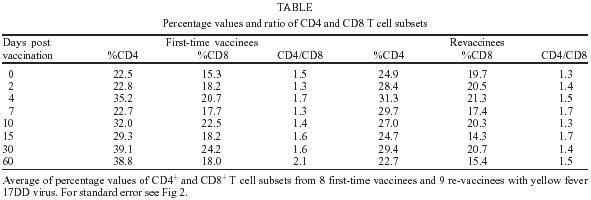
2004 Code number: oc05093 In this study the kinetics of humoral and cellular immune responses in first-time vaccinees and re-vaccinees with the yellow fever 17DD vaccine virus was analyzed. Flow cytometric analyses were used to determine percentual values of T and B cells in parallel to the yellow fever neutralizing antibody production. All lymphocyte subsets analyzed were augmented around the 30th post vaccination day, both for first-time vaccinees and re-vaccinees. CD3+ T cells increased from 30.8% (SE ± 4%) to 61.15% (SE ± 4.2%), CD4+ T cells from 22.4% (SE ± 3.6%) to 39.17% (SE ± 2%) with 43% of these cells corresponding to CD4+CD45RO+ T cells, CD8+ T cells from 15.2% (SE ± 2.9%) to 27% (SE ± 3%) with 70% corresponding to CD8+CD45RO+ T cells in first-time vaccinees. In re-vaccinees, the CD3+ T cells increased from 50.7% (SE ± 3%) to 80% (SE ± 2.3%), CD4+ T cells from 24.9% (SE ± 1.4%) to 40% (SE ± 3%) presenting a percentage of 95% CD4+CD45RO+ T cells, CD8+ T cells from 19.7% (SE ± 1.8%) to 25% (SE ± 2%). Among CD8+CD38+ T cells there could be observed an increase from 15 to 41.6% in first-time vaccinees and 20.7 to 62.6% in re-vaccinees. Regarding neutralizing antibodies, the re-vaccinees presented high titers even before re-vaccination. The levels of neutralizing antibodies of first-time vaccinees were similar to those presented by re-vaccinees at day 30 after vaccination, indicating the success of primary vaccination. Our data provide a basis for further studies on immunological behavior of the YF 17DD vaccine. Key words: yellow fever - vaccine - flow cytometry - neutralizing antibodies - lymphocyte subsets Yellow fever (YF) is a viral illness transmitted by infected mosquitoes (Aedes and Haemagogus genus) with yellow fever virus (YFV), which belongs to the Flavivirus genus, Flaviviridae family (Monath & Heinz 1996). It remains a serious health problem in endemic areas of tropical and subtropical Africa and South America (Vasconcelos 2003). Since the 19th century, studies about YF immunization have been accomplished (WHO 2001a). The YF 17D vaccine virus strain is one of the most effective and safe vaccines available. Immunization with the 17D vaccine strain induces a long-term neutralizing antibody response and provides excellent protection against infection with the virulent YFV (Wisseman & Sweet 1962). In 1937, the production of the YF 17DD vaccine virus substrain was started in Brazil, and has been in continuous use for 60 years (Post et al. 2001). The YFV 17DD vaccine is highly immunogenic and induces neutralizing antibody persistent at least 10 years and in some individuals up to 30 years or more (WHO 2001b). Although the YF 17D virus strain and the 17DD substrain are the most successful vaccines developed to date, recently, rare cases of postvaccinal neurological disorders have been recorded (Chan et al. 2001, Martin et al. 2001, Vasconcelos et al. 2001). The virus isolates from two fatal cases after 17DD vaccination demonstrated genetic stability and attenuated phenotype, suggesting that some peculiarities of health status of the host might have been responsible for such adverse events (Galler et al. 2001). Despite the large literature on the humoral immune response to the YF vaccine virus, few works of the evaluation of cellular immune response, in particular T cell responses have been published (Reinhardt et al. 1998, Co et al. 2002, van der Most et al. 2002). The capacity of vaccines to activate the cellular immune system and induce T cell memory is an important mechanism of protection against wild-type viral infection. T cells often recognize more conserved epitopes that do not change due to antibody mediated selection pressure (Whitton & Oldstone 1996, Reinahrdt et al. 1998). Co and collaborators (2002) studying the T cell responses to YFV 17D in four volunteers could observed proliferation and cytolytic responses in all subjects. Their results present CD8 T cell responses directed against at least four different HLA-B35 restricted YFV epitopes. In order to monitor the immunological behavior of first-time YF 17DD virus vaccinated and re-vaccinated volunteers, T and B lymphocyte subsets were analyzed by flow cytometry, which is an efficient tool for definition and quantification of lymphocytes employing monoclonal antibodies against cell surface proteins, to determine the importance of knowing the cellular immune response activation related to the YF 17DD vaccine. MATERIALS AND METHODS Volunteers - Seventeen healthy persons with 18 to 55 years of age were vaccinated with the YF 17DD vaccine (Lot 007VFA010Z), produced in Bio-Manguinhos, Oswaldo Cruz Foundation, Brazil. Eight individuals, designated first-time vaccinees, had not been vaccinated with the YF 17DD vaccine, and had had no previous infection or contact with the YF wild-type virus. Nine volunteers, designated re-vaccinees, had been vaccinated once or more with the YF 17DD vaccine, 10 or more years before. All volunteers gave their written consent after the study was explained. Each volunteer was injected subcutaneously with 0.5 ml of vaccine. Blood was collected before vaccination (day 0) and 2, 4, 7, 10, 15, 30, and 60 days post vaccination. The volunteers were advised to report all clinical symptoms and side effects after vaccination. The procedures followed were in accordance with the ethical standards of the responsible committee on human experimentation (CEP/Fiocruz 145/01). Separation of peripheral blood mononuclear cells (PBMC) - PBMC were obtained by Histopaque (Sigma-H8889, St Louis, MS, US) gradient sedimentation, as described elsewhere (Noble et al. 1968, Boyle & Chow 1969). Flow cytometric analyses - Freshly isolated PBMC were adjusted to 106 cells/ml and dual-staining labeled monoclonal antibodies (CD3(IgG1)-FITC/B4-RD1; CD4-PC5/ CD45RO-PE; CD8-PC5/CD45RO-PE; CD8-PE/CD38-FITC) and IgG1-FITC/IgG1-PE isotype antibody (negative control) (Immunotech - Beckmann Coulter, Marseille, France) were used. After incubation for 20 min at 4ºC, the mixture was washed twice with PBS-azide (PBS containing 0.1% sodium azide and 2% fetal bovine serum), and the cells were fixed in 1% paraformaldehyde (PFA). Stained cells were run in an EPICS ALTRA flow cytometer (Beckmann Coulter, Hialeah, FL, US) equipped with an argon ion laser. Ten thousand events were acquired and analyzed using the Expo32 software (Beckman Coulter). Their forward scatter and side scatter profiles distinguish lymphocytes and monocytes, an electronic gate was created around lymphocytes. Plaque reduction neutralization test (PRNT) - PRNT were carried out in 96-well plates as described elsewhere (Stefano et al. 1999). In brief, dilutions of the sera ranging from 1:4 to 1:512 were mixed with 1:1200 of the 17DD virus preparation (7.8 log10 PFU/ml). After 1 h of incubation at 37oC, 100 µl of Vero cells (1.6 x 106 cells/ml) were added. A 3% CMC/199 solution supplemented with 5% FCS was overlaid after 3 h of incubation. The cultures were maintained for 7 days at 37oC and then the cells were fixed with formaldehyde and stained by violet crystal. Viremia - For determination of viremia were used the plaque assay (PA) (Wheelock & Sibley 1965, Reinhardt et al. 1998, Marchevsky et al. 2003) and the reverse transcriptase-polymerase chain reaction (RT-PCR) (Reinhardt et al. 1998) methods, in serum samples before vaccination and on days 2, 4, and 7 after vaccination. RT-PCR products were obtained in the presence of pairs of flavivirus type-specific primers (RG65-antisense, RG36-sense) (Galler et al. 2001), which were kindly provided by Dr Ricardo Galler (Oswaldo Cruz Institute - Fiocruz). Primer pairs were deduced from a RNA sequence from the envelope coding region of the yellow fever virus. The RT-PCR techniques were performed using the Geneamp kit (Applied Biosystems RNA PCR Core Kit, Roche, Branchburg, New Jersey, US) according to the manufacturer's instructions. Statistical analyses - Fisher's least significant difference (LSD) procedure was used to evaluate statistically significant differences between means. With this method, there is a 5% risk of calling each pair of means significantly different when the actual difference equals 0. Differences were considered as statistically significant if p value were equal or below 0.05. RESULTS The 17DD vaccine was shown to be very efficient and safe for all volunteers, activating their humoral and cellular immune response, although viremia was not detected through of the methods used (RT-PCR and PA). During the kinetics blood collection in this study, the blood cell count and biochemical tests, including transaminases, did not present alterations after vaccination for any of the volunteers. None of the first-time vaccinees showed pre-existing neutralizing antibodies (nAb) against the YF 17DD virus substrain (< 256 mUI/ml, limit of antibody detection for our test). Four days after vaccination, low levels of nAb were detected in 3 of the 8 first-time vaccinees (mean 698 mUI/ml, SE ± 301), however, 15 days after vaccination, the nAb were detected in all of them. High levels of nAb were reached 30 days after vaccination in this group of volunteers with a slight decrease 30 days later, 30,321 mUI/ml (SE ± 1300) and 22,756 mUI/ml (SE ± 4126), respectively. All re-vaccinees had persistent nAb before re-vaccination, with a mean titer of 5840 mUI/ml (SE ± 2431), despite the interval of, at least, 10 years since the last vaccination. The high standard error achieved was due to the high nAb titer of one volunteer, above 14,000 mUI/ml, before revaccination. After re-vaccination, nAb levels increased to a mean titer of 28,526 mUI/ml (SE ± 2691) and 21,598 mUI/ml (SE ± 3821) 30 and 60 days later, respectively. Only one volunteer showed no changes in nAb levels. The induction of nAb was significantly different between both groups studied among day 0 and day 15 after vaccination (p < 0.05) (Fig. 1). Figs 2, 3, and 4 show the T cell subsets from volunteers vaccinated with the YF 17DD virus. In all volunteers an increase in CD3+ T cells was observed. In first-time vaccinees, the level of circulating CD3+ T cells has significantly increased, from 30.8% (SE ± 4.0%) of the mononuclear cells before vaccination to 61.15% (SE ± 4.2%) on the 30th day post vaccination (p = 0.0002) and in re-vaccinees from 50.7% (SE ± 3.0%) to 66.5% (SE ± 2.6%) on the 7th day for 6 of the 9 volunteers (p = 0.0001) and 80% (SE ± 2.3%) on the 30th day after vaccination in the other volunteers (n = 3) (p = 0.0464). In first-time vaccinees a significant increase in the levels of circulating CD8+ T cells was observed. The initial mean in first-time vaccinees increased from 15.2% (SE ± 2.9) to 27% (SE ± 3.0%) on the 30th day post vaccination (p = 0.0276), with 70% of the cells being identified as memory cells, according to the analysis of the CD8+ CD45RO+ T cell subset. Among re-vaccinees, the initial concentration of these cells was 19% (SE ± 1.8%) before vaccination and 22.5% (SE ± 2.0%) in 5 volunteers between the 4th and the 10th day (p = 0.2499) and 25% (SE ± 2.0%) on the 30th day post vaccination in the four other volunteers (p = 0.0083) with identification of 90% of the cells as being CD8+CD45RO+ T cells in all volunteers, at day 30. Among the CD8+CD38+ T cells, a marker for activated T cells, a significant increase could be observed, from 15 to 41.6% in first-time vaccinees (p = 0.0207) and from 20.7 to 62.6% in re-vaccinees (p = 0.0017). Analyzing the levels of CD4+ T cells, it could be observed that a significant increase in first-time vaccinees occurred, from 22.4% (SE ± 3.6) to 39.17% (SE ± 2.0%) on the 30th day post vaccination (p = 0.0026), with 43% of these cells being identified as CD4+CD45RO+ T cells. The re-vaccinees have presented levels of circulating CD4+ T cells from 24.9% (SE ± 1.4%) to 35.24% (SE ± 1.2%) between the 4th and the 10th day (n = 5) (p = 0.1613), and to 40% (SE ± 3.0%) on the 30th day post vaccination in the four other volunteers (p = 0.0051), presenting a percentage of 90 and 95% of CD4+CD45RO+ T cells, respectively. The CD4/CD8 ratios were above one (varying from 1.3 to 2.1) with no difference between the two groups studied (Table). DISCUSSION The YF 17DD vaccine, produced in Bio-Manguinhos-Fiocruz, Brazil, is known to be a safe and efficient vaccine, leading to suitable results with regards to nAb production (Stefano et al. 1999, WHO 2001b, Marchevsky et al. 2003). However, the cellular immune response profile of the YF 17DD vaccine has not been known as yet, although some rare adverse events after vaccination have been suggested as a failure in the cellular immune response of these patients (Galler et al. 2001, Mariannaeu et al. 2001, Vasconcelos et al. 2001). In this work, we have analyzed some parameters of humoral and cellular immune responses after YF 17DD vaccination. The volunteers of this study have well tolerated the vaccination. No significant changes were detected in red and white blood cell and platelet counts or in the transaminase levels after vaccination. The methods used to monitor the viremia, PA and RT-PCR, were not able to detect viruses or viral RNA fragments in the sera of the first-time vaccinees or re-vaccinees, despite of these methods were used with success by other authors (Reinhardt et al. 1998). A hypothesis for this lack of detection of viremia could be the high titers of neutralizing antibodies, which would be able to suppress the presence of circulating virus (Wisseman Jr & Sweet 1962) and the decrease of viral circulation that is prompted by YF 17DD virus, unlike the wild type virus (Monath 1998). In a previous study, the RT-PCR was shown to be more sensitive than PA, it was seen that the RT-PCR detects stretches of virus-specific RNA that may represent only RNA fragments or defective viral particles, as a positive assay does not necessarily reflect the presence of viable virus (Reinhardt et al. 1998). All re-vaccinees have presented high titers of persistent neutralizing antibodies before re-vaccination, although they have not been in yellow fever endemic areas within ten years of re-vaccination. These results strengthen the idea of viral presence in lymphatic tissues suggested by other reports (Reeves et al. 1958, Oehen et al. 1992). The titers of nAb in all re-vaccinees have reached high values on the 30th day after revaccination. The induction of nAb observed among the first-time vaccinees have indicated the success of the vaccination. The levels of nAb in this group of volunteers were similar to those shown for the re-vaccinees. The CD3+ T cell levels were analyzed to evaluate the profile of cellular immune responses in all vaccinated subjects. All volunteers have presented a significant increase in his T cell subset, suggesting an activation of the cellular immune response. With respect to CD8+ T cell counts, it was observed a significant augmentation in all first-time vaccinees and in four re-vaccinees volunteers. This increase of CD8+ T cells was expected due to the importance of these cells in the mechanism of the immune response for intracellular attenuated pathogens vaccinees described by other authors (van der Most et al. 2000, 2002, Co et al. 2002). In our study, the vaccination induced the activation of CD8+ T cells, as could be observed on the significant increase of the CD8+CD38+ T cells in all volunteers. CD4+ T cells are involved in several steps in the mechanics of immune response, as antibodies and cytokine production, among other events during the type-2 immune response, through B cells activation and posterior plasma cell differentiation (Mills et al. 2000, Liu & Chambers 2001). As was observed in this study, all first-time vaccinees have presented a significant increase in their numbers of B cells (data not shown) and in the CD4+ T cell subsets. In the group of the re-vaccinees, only four volunteers have presented a significant increase in the latter lymphocyte subset. It is important to note that these volunteers also had their CD8+ T cells augmented significantly, as discussed above. The CD4/CD8 ratio did not demonstrate difference between the two analyzed groups. The number of CD4+ T cells were more than CD8+ T cells, showing a normal activation of the immune system after vaccination, to the opposite of the infection with flavivirus which presents CD4/CD8 ratio inversion (Lei et al. 2001, Liu et al. 2002). Not all re-vaccinees have presented significant increase in the levels of CD4+ and CD8+ T cells, but the percentage of their activated cell subsets (CD4+CD45RO+ and CD8+CD45RO+ cell markers) was more highly augmented than those of the first-time vaccinees. This result has suggested that the re-vaccinees were able to answer to the YF 17DD virus without need for a co-stimulation. The importance of the CD4+ T cell and CD8+ T cell subsets in nAb production could be observed through analysis of the results from one of the volunteers with leucopoenia, anemia and thrombocytopenia before vaccination. In this volunteer, the increase of the cell subsets analyzed did not occur and the nAb induction was observed later than in the other volunteers. Despite of several studies carried out analyzing the levels of lymphocyte subset standards in healthy humans, it is known that such values are not constant, some factors can affect lymphocyte subset counts, such as stress, physical exercises, smoking and alcohol (Goff et al. 1985, Uppal et al. 2003). The data presented herein provide a basis for further studies on immunological response to the YF 17DD vaccine in human beings. Therefore, a study of the T cell immune response activation associated to lymphocyte proliferation and cytokine production in YF vaccination is underway. ACKNOWLEDGMENTS To Centro de Pesquisa Hospital Evandro Chagas-Fiocruz for measuring blood cells counts and serum biochemical assays and the technical assistance of Adriana de Oliveira Honorato, Bio-Manguinhos-Fiocruz. To Dr Vera Bongertz from De-partamento de Imunologia, Instituto Oswaldo Cruz-Fiocruz for reading the manuscript. REFERENCES
The following images related to this document are available:Photo images[oc05093t1.jpg] [oc05093f4.jpg] [oc05093f1.jpg] [oc05093f2.jpg] [oc05093f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}