 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Memórias do Instituto Oswaldo Cruz, Vol. 100, No. 4, July 2005, pp. 407-420 Angiostrongylus costaricensis: complete redescription of the migratory pathways based on experimental Sigmodon hispidus infection Ester Maria Mota+, Henrique Leonel Lenzi Departamento de
Patologia, Instituto Oswaldo Cruz-Fiocruz, Av. Brasil 4365, 21040-900 Rio
de Janeiro, RJ, Brasil Received 31 August
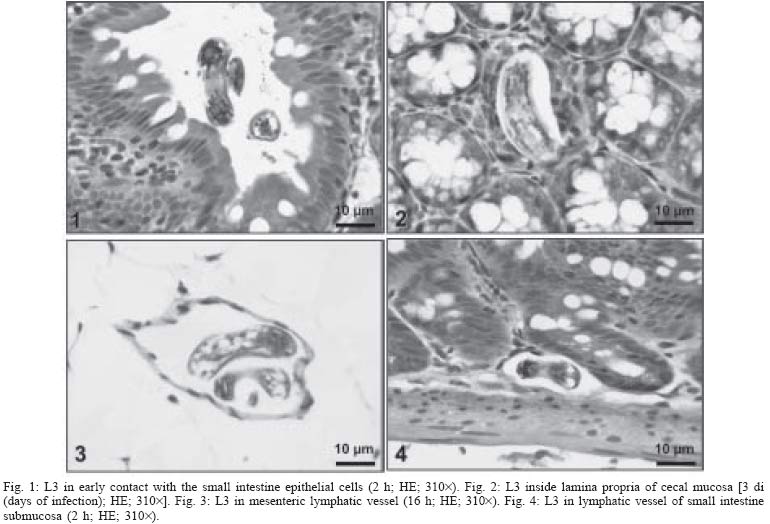
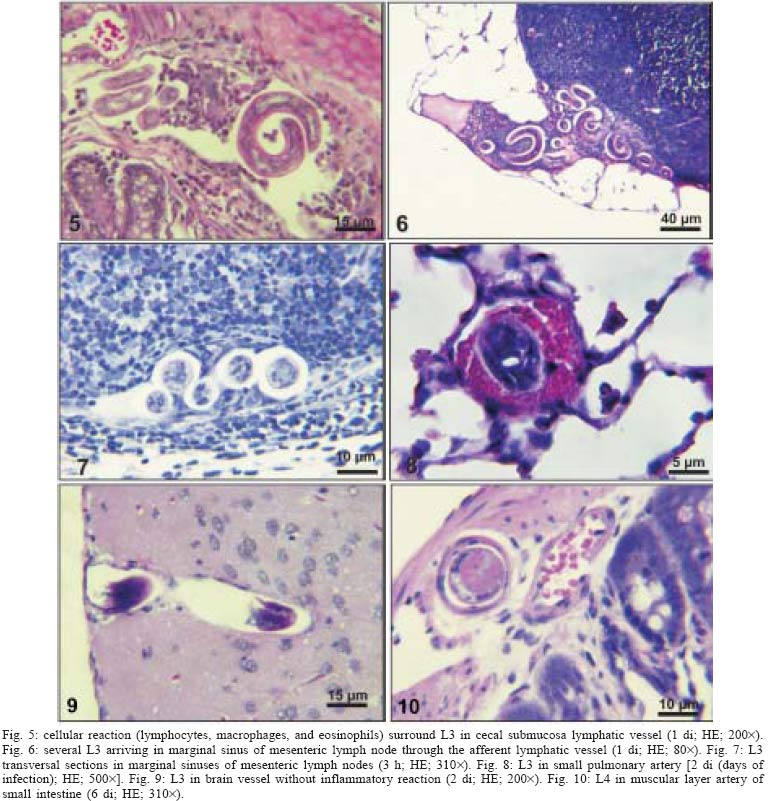
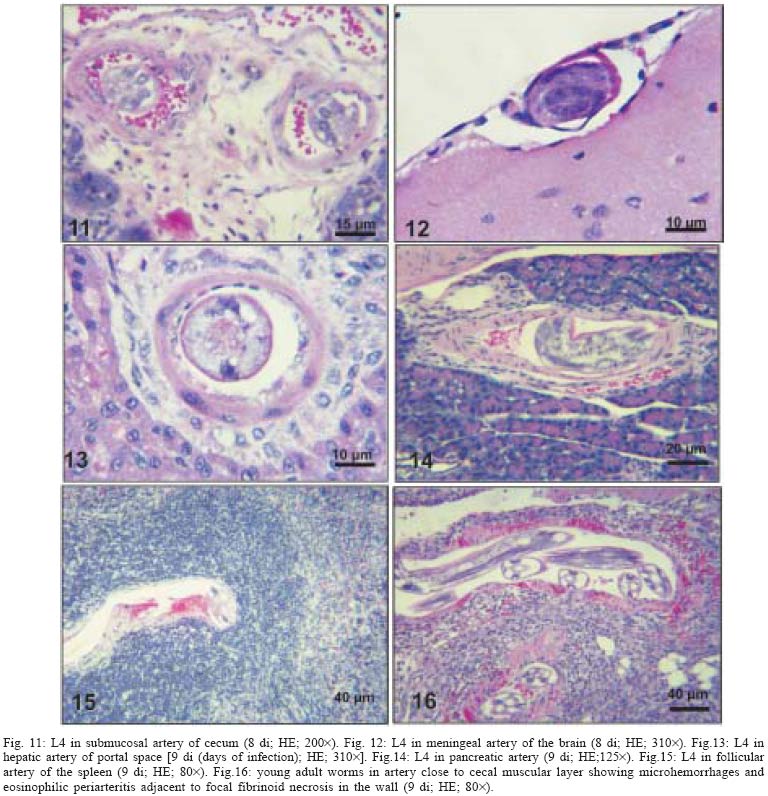
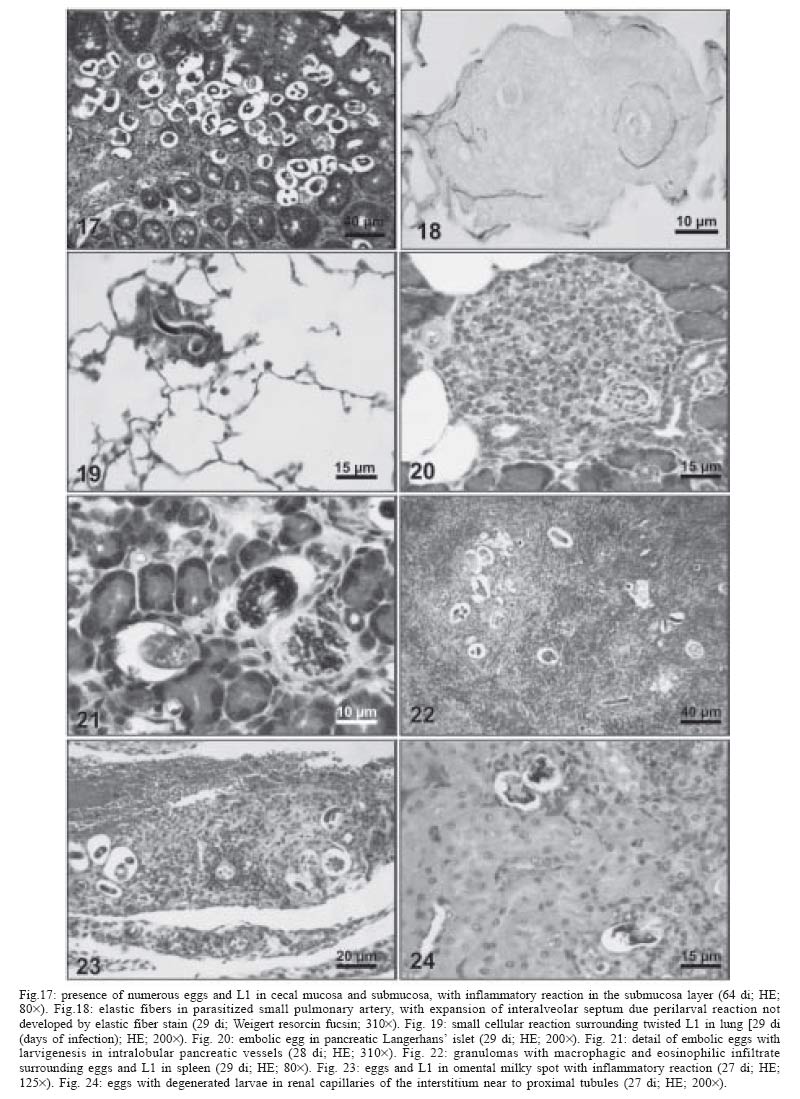
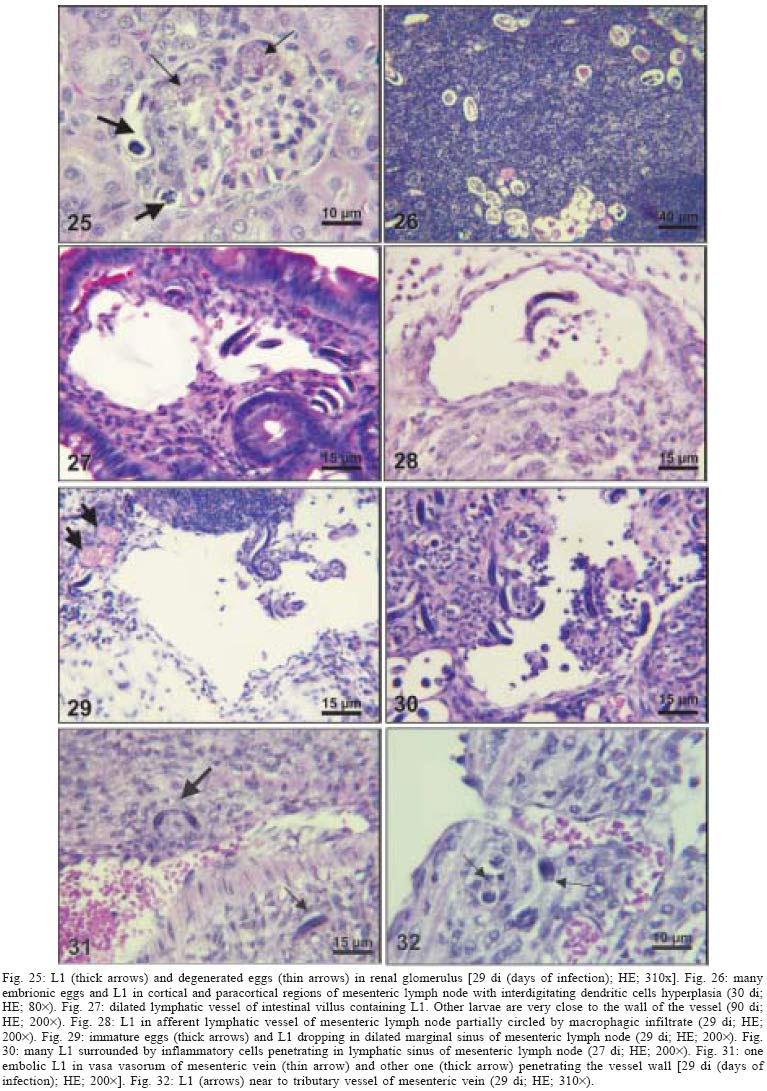
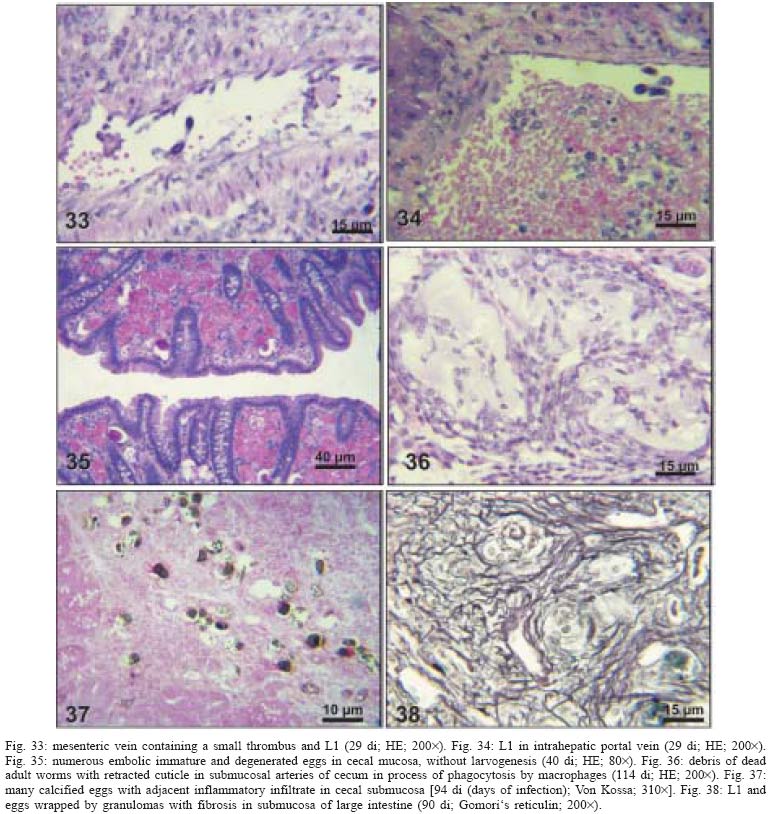
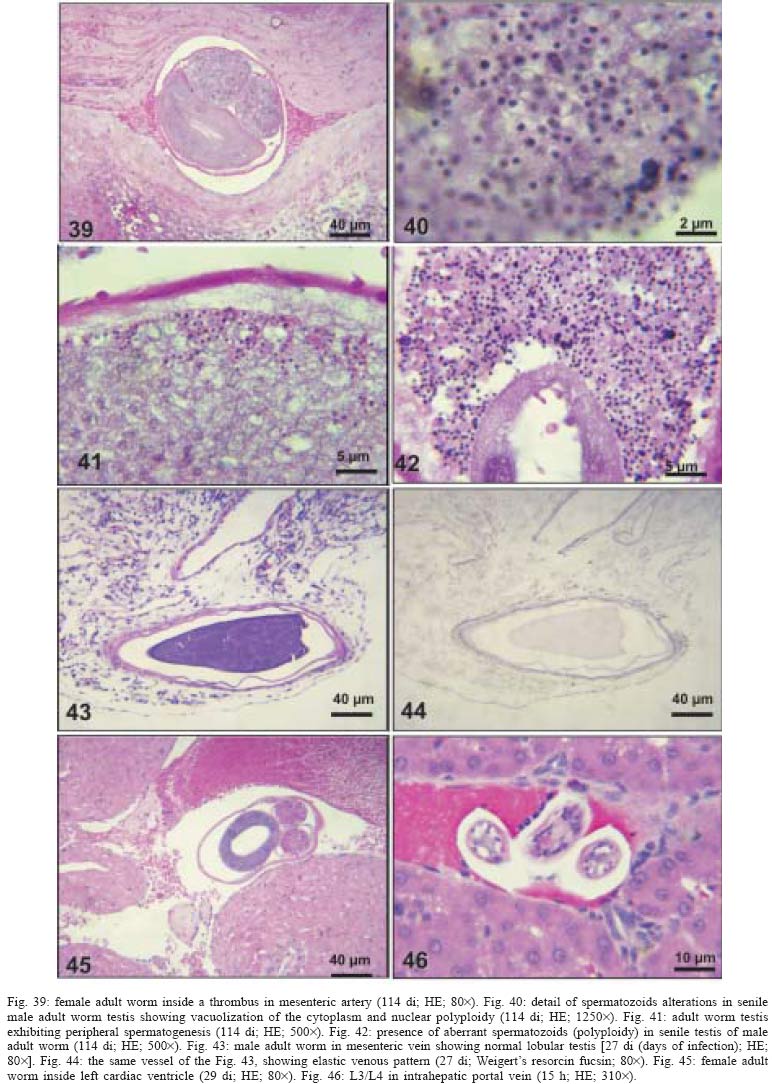
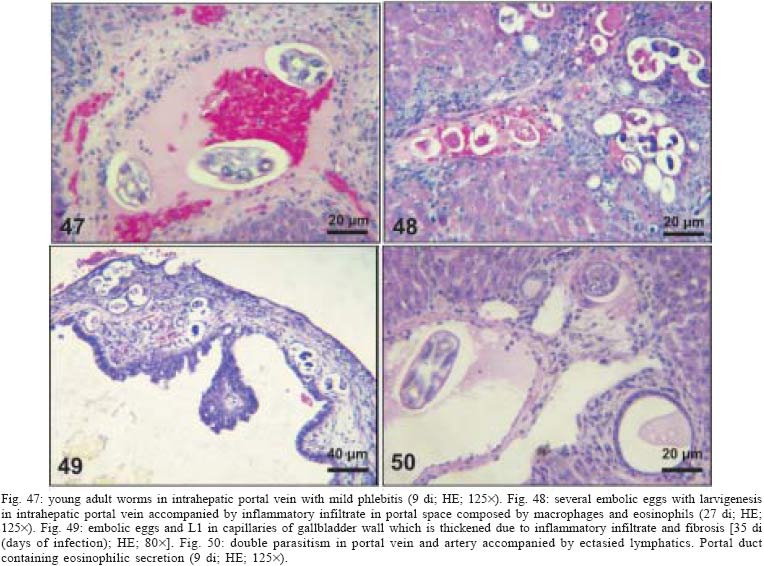
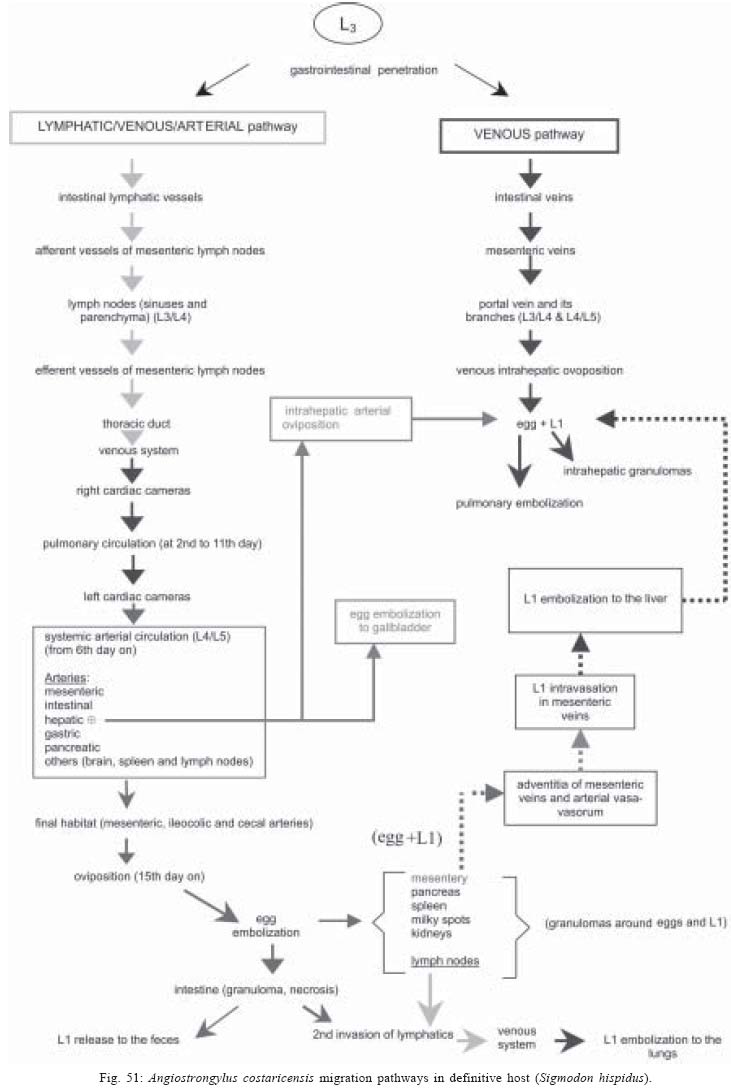
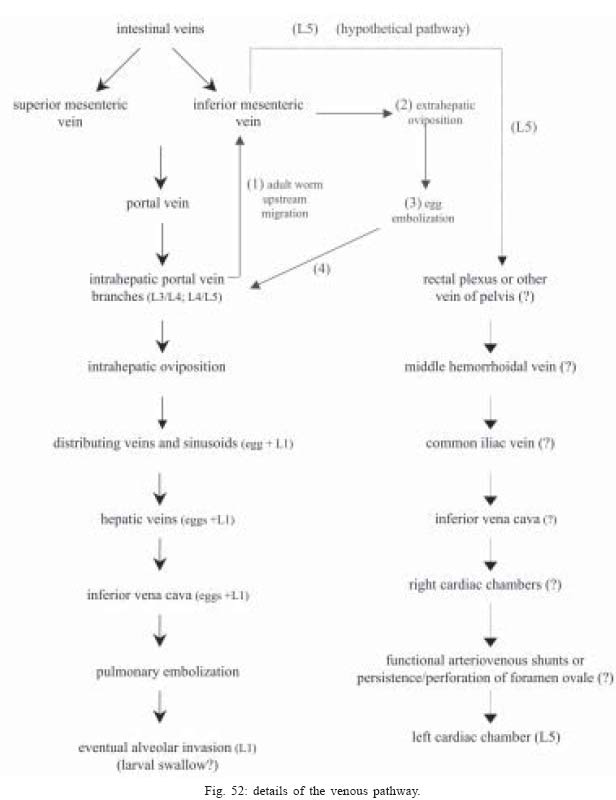
2004 Code Number: oc05105 Angiostrongylus costaricensis lives in the cecal and mesenteric arteries of its vertebrate hosts, and causes an inflammatory disease in humans. To investigate unknown aspects of the abdominal angiostrogyliasis pathogenesis, infected Sigmodon hispidus were sequentially studied in different times of infection. The study revealed that L3 goes alternatively through two migratory courses during its development into an adult worm: lymphatic/venous-arterial and venous portal pathways. The former is considered the principal one, because it is used by most of the larvae. Like other metastrongylides, A. costaricensis passes over the pulmonary circulation to migrate from the lymphatic system to the arterial circulation, where they circulate during some days before reaching their definitive habitat. The oviposition by mature females began on 15th day. Eggs and L1 were detected mainly in the intestine and stomach, surrounded by inflammatory reaction constituted by macrophages, monocytes, and eosinophils. They were also spread to the lungs, mesenteric lymph nodes, pancreas, spleen, and kidneys. The larvae (L1) exhibited the centripetal capacity to invade the lymphatic and venous vessels of the intestine and mesentery. Adult worms that developed in the venous intrahepatic pathway migrated downstream to reach the mesenteric veins and laid eggs that embolized in the portal hepatic vessels. Key words: Angiostrongylus costaricensis - life cycle - Sigmodon hispidus - liver Angiostrongylus costaricensis is a Metastrongylidae worm that causes a human parasitism named abdominal angiostrongyliasis (Morera & Céspedes 1971). This disease is characterized by the massive infiltration of eosinophils in the intestinal wall, granulomatous reaction, and eosinophilic vasculitis affecting arteries, veins, lymphatics, and capillaries, mainly in the ileo-cecal region (Céspedes et al. 1967, Graeff-Teixeira et al. 1991). This helminth is found from the United States to Argentina, but most of the cases occur in Central America. In Brazil, this zoonosis was detected in São Paulo (Zillioto et al. 1975), Distrito Federal (Barbosa et al. 1980), Minas Gerais (Rocha et al. 1991), and Espírito Santo (Pena et al. 1995), but the Southern region is the predominant endemic area (Agostini et al. 1983, Ayala 1987). Human beings are considered accidental hosts because they do not eliminate larvae in the feces. Men acquire the parasite by the ingestion of L3 on uncooked vegetables or fruits. There is no specific treatment to human patients because usual anti-helminthic drugs are not effective and may worsen the clinical picture (Morera & Bontempo 1985). However, in mice models, treatment with mebendazol was not associated with distal migration of adult (Mentz et al. 2004) and inhibited egg formation and/or oviposition of A. costaricensis (Terada et al. 1993). Ishii et al. (1989) observed that levamisole had conspicuous in vivo effects against larval stages of A. costaricensis in mice. A. costaricensis mature adults live in the branches of the superior mesenteric artery of rats and other rodents. The females lay eggs that are taken by the blood current in the intestinal wall where larvogenesis occurs generating L1. They hatch and are eliminated with the feces. In nature, these larvae can penetrate intermediate hosts (mollusk) through oral (Morera 1973) and/or percutaneous infections (Thiengo 1996, Mendonça et al. 1999). In the muscular layer of the invertebrate host they suffer two moultings (L1→ L2→ L3) and further elimination of L3 within the mucous secretion (Conejo & Morera 1988, Mendonça et al. 1999). Rodents are probably infected by eating molluks or vegetables contaminated by L3. The A. costaricensis evolution in the definitive host, Sigmodon hispidus, was first described by Morera (1973). Actually, in this classical paper, the author described the complete cycle in the mollusk and in mammalian. According to this author, in the rodent the cycle takes place entirely in the abdominal cavity, and the young adults migrate to their definitive habitat by moving from the lymphatic vessels to the mesenteric arteries through the vascular walls provoking multiple small hemorrhages around the vessels. However, several reports on naturally infected animals and patients have described the occurrence of different parasitic stages (eggs, L1, and adult worms) in other organs not mentioned in the original description of the life history of A. costaricensis (Velazquez et al. 1974, Morera et al. 1982, Duarte et al. 1991, Vazquez et al. 1994, Brack & Schropel 1995, Zuccaro et al. 1998). In order to elucidate these findings on migratory pathways of A. costaricensis, Mota and Lenzi (1995) have proposed a new conception of this nematode life cycle based on a mouse model similar to the other Metastrongylides. They showed that the nematode passage from the lymphatic to the arterial vessels occurs through the pulmonary circulation and they described two migratory routes: a lymphatic-venous/arterial and a venous portal pathways. The present report gives a complete redescription of the life history of A. costaricensis based on the experimentally infected S. hispidus, showing that the findings are very similar to those described in the mouse model. The study used S. hispidus (cotton rat) because it is the main definitive host in the wild cycle in Central America (Morera 1970, Morera et al. 1977). MATERIALS AND METHODS Parasite and infection - A. costaricensis (obtained from Crissiumal, RS, Brazil) life cycle has been maintained in laboratory using cotton field rats and Sarasinula marginata (Semper, 1885) slugs. The third stage larvae were obtained by the sacrifice of infected mollusks. The muscular tissue was cleaved and digested for two hours at 37ºC by the Wallace and Rosen (1969) method. The infectious larvae were inoculated in cotton field rats by using a gastric probe. As of 28 days on, the feces of infected rodents were collected and put in a Baermman apparatus to separate from the decanted L1. These larvae were given to the slugs with pieces of lettuce. Vertebrate host - A colony of cotton field rats S. hispidus (imported from Virion System Inc., Rockville, MD, US) is bred in the Pathology Department, Instituto Oswaldo Cruz-Fiocruz, Rio de Janeiro. Experimental design - Two hundred sixteen rodents at three months of age (both sexes) were divided into groups of eight animals and orally infected with 100 and 500 L3 at each infection, except at 2, 3, and 16 h, when 4000 larvae were injected. The study was performed at 2, 3, and 16 h and on days 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 13, 15, 27, 28, 29, 30, 35, 40, 50, 60, 80, 90, and 114 after the infection. Animals were submitted to euthanasia by ether inhalation in accordance with the animal ethical practice. The organs (brain, thymus, heart, lungs, esophagus, stomach, intestine, mesentery, liver, pancreas, spleen, kidney, adrenal glands, and genital apparatus) were fixed in Carson's Millonig-formalin (Carson et al. 1973) and embedded in paraffin. Serial sections (5 µm) were consecutevely stained with Hematoxylin and Eosin; Lennert's Giemsa (Lennert 1978) for cells; Weigert's resorcin-fucsin for elastic fibers; Masson's trichromic for collagen fibers; sirius red pH 10.2 for eosinophil (Bogolometz 1980, Luque & Montes 1989), Von Kossa for calcium deposits and Perls' Prussian blue reaction for ferric iron. The images were captured with a Hamamatsu color chilled 3CCD camera, (model C-5810) and recorded in Microsoft Photo Editor. RESULTS Two hours after the infection, L3 were only observed in gastric lumen and close to the epithelium of the small intestine villosities (Fig. 1). From 3 h to 11 days after the infection, the larvae were inside the lymphatic vessels of the stomach, small and large intestines, mesentery, and pancreas (Figs 2, 3). The larvae preferentially occupied the lymphatic vessels of submucosa (Figs 4, 5), including Peyer patch, but were also present in the mucosa, muscular and serosa layers of the intestine. Many larvae were also detected in afferent and efferent lymphatic vessels and in marginal sinuses and cortical region of mesenteric lymph nodes (Figs 6, 7). The cecum and large intestine were the main sites of penetration and lymphatic invasion by the parasites. The presence of parasites provoked an inflammatory reaction in the abdominal lymphatic circulation, evidenced by lymphangitis and peri-lymphangitis constituted by macrophages, eosinophils, and neutrophils. During the passage of larvae in the abdominal lymphatic system, they suffered molt to L4 increasing the length and sketching the reproductive organ. On 11th day, in the efferent lymphatic vessel and in the mesenteric lymph node, few damaged L4 surrounded by eosinophils in the center of a granuloma were noticed. On day two of infection, an L3 was observed inside a distal pulmonary artery close to pulmonary alveoli (Fig. 8) and another one was in the meningeal vessel (Fig. 9). From 6th day on, groups of L4 were also present in the cecal and large intestine muscular and submucosa arteries (Figs 10, 11). The number of parasites in the arterial vessels increased from day 9 onward and they began to appear also in the mesentery, small intestine, stomach, liver, spleen, pancreas, brain, and liver (Figs 12-15). Parasitism in the arterial system was accompanied by increasing of eosinophil and macrophage infiltration and the arising of eosinophilic periarteritis with or without fibrosis, fibrinoid necrosis of the muscular layer, and microhemor-rhages in the arterial wall (Fig. 16). In arterial vessels, the L4 suffered the last molt and became young adult worms with the maturation of reproductive organs. On 11th day, most of the parasites reached their definitive habitat in the arterial vessels of the cecum, large intestine, and mesentery. Many immature and embrionated eggs were carried by the blood current, like emboli, and impacted in the mucosa layer of the small and large intestines, mainly in the cecum on 15th day. From 27th day on, many L1 and eggs were present in all layers of the intestinal and gastric walls surrounded by inflammatory reaction constituted essentially by macrophages, but monocytes and many eosinophils also participated (Fig. 17). Eggs in different stages of embriogenesis and L1 were also embolized in the lungs, mesentery, mesenteric lymph nodes, pancreas, spleen, omental milky spots, and kidneys (Figs 18-26). With the exception of the kidney, the eggs and L1 did not present morphological changes. Some L1 left the intestinal mucosa by active moving and/or were expelled with necrotic detritus, together with the eggs, in mucosa ulcerations. Sometimes, larvae and eggs were inside the ectatic lymphatic vessels of the large and small intestine mucosa (Fig. 27). These parasitic forms were transported by the lymph, reaching the mesenteric lymph nodes (Fig. 28). Lymphatic sinuses of lymph nodes were also invaded by L1 originated from local larvogenesis (Figs 29, 30). L1 also migrated to the vessels around the mesenteric veins, and penetrated inside the vascular wall falling into the lumen. This event seemed to be facilitated by the inflammatory infiltration that injured the venous wall and destroyed the elastic fibers of the vessel (Figs 31-33). Some of the intravascular L1 could reach the intrahepatic vessels by upstream transportation (Fig. 34). From 30 days on, the presence of degenerated eggs and intra-arterial dead adult worms became constant (Figs 35, 36). On day 80 of the infection, many calcified and infertile eggs were retained in the cecal mucosa (Fig. 37). On 90th day, few L1 living inside fibrous cocoons were immobilized in granulomas (Fig. 38). On day 114, in spite of the larvogenesis absence, there were few living males and females inside the arteries, most of them next to the thrombus (Fig. 39). The testis of some males presented vacuolated and anomalous spermatozoids (Figs 40-42). Adult worms were always present inside the mesenteric and intestinal arteries. They were detected in the mesenteric lymph nodes and pancreatic arteries until 28 days after the infection. An adult worm, eggs and L1 were present in the mesenteric vein on 27th day (Figs 43, 44). Curiously, in two animals, with 29 and 30 days of infection, adult worms were found in the left cardiac ventricle, being male in one case and female in the other one (Fig. 45). The development and maturation of A. costaricensis in the liver occurred together and independently of the parasites present in the lymphatic or arterial circulation. The L3 arrived in the venous intrahepatic vessels 3 h after inoculation, and there suffered two molts to adult worms, laying eggs locally (Fig. 46). L1 and eggs could be found in portal veins (conducting and distributing veins) and/or sinusoids, surrounded or not by inflammatory cells, but most of them were in small vascular branches (distributing vessels) around portal spaces. Venous intrahepatic adult worms were detected until 9 days after the infection (Fig. 47), although eggs and L1 were embolized in intra-hepatic venous vessels until the end of the experiment (Fig. 48). Embolic eggs were also detected in vessels of the gallbladder wall (Fig. 49). Young and mature adult worms were in space portal arteries from 8th until 27th day (Fig. 50). The lack of larvogenesis inside the liver occurred simultaneously with the same event in the intestinal wall. DISCUSSION On this study we showed that the life history of A. costaricensis in its natural vertebrate host (S. hispidus) is much more complex than originally described by Morera (1973). It reproduces, with some individual variation, the data observed in the mouse model, and explains better the findings reported in natural infected animals and patients, such as the venous migration to the liver. In fact, like other metastrongylides, A. costaricensis uses the pulmonary circulation to pass from the lymphatic/venous to the arterial or systemic circulation. In spite of the frequent occurrence of fibrinoid necrosis and multiple microhemorrhages in arteries from different regions, during the development of L3 into an adult worm (before 15 days of infection), L4/L5 extravasation was never observed. Only L1 showed great capacity of tissue migration, and alone or together with surrounded inflammatory cells, they can reinvade the lymphatic system (secondary or late invasion of lymphatics) pervading the intestinal lymphatics and lymph node sinuses, or even, they can go into mesenteric veins, departing from perivenous embolization foci (intravasation). This phenomenon can give rise to secondary "metastatic" embolization of L1 to intrahepatic venous portal branches. This original event, for the first time described in nematodes is different from the autoinfection caused by Strongyloides stercoralis L3, when L1 have time to molt twice during their transit down the digestive tract, acquiring capacity to penetrate the skin, go through their migration, and mature (Roberts & Janovy Jr. 1996). The A. costaricensis, in spite of its preference for arterial vessels, can also live for a while inside the venous system (L4/L5) and lymphatic vessels (L1, L3/L4), indicating its physiologic ubiquitous capacity of living in microenvironments with different O2 tension. It is metabolically flexible like other nematodes, combining the versatility of an anaerobic (L1, L3/L4) and aerobic energy metabolism (L4/L5) (Fry & Jenkins 1984). The adult worms seem to be mandatory aerobes with respect to their energy metabolism, requiring the presence of at least low oxygen concentration (venous blood) and red blood cells (RBC) for survival and mobility. The addition of RBC to Waymouth's medium was essential for A. costaricensis further development, although it was not essential for the development itself up to the young adult stage (Hata & Kojima 1991). Probably, the oxygen suppression in a vessel with thrombosis can explain the death of adult worms trapped inside the thrombus. This occurrence can also place living males and females at a distance, avoiding sexual contact, promoting consequently a large quantity of infertile eggs, observed in some infected animal, mainly in the more advanced stages of the infection. The female sterility can be an effect of male aging, expressed by anomalous spermatozoids in the testis. The reason for the metabolic or biochemical preference of L3 to intestinal and mesenteric lymphatic vessels, where they suffer molt to L4 is unknown. Perhaps they utilize the water-insoluble fats absorbed from the intestine into the lymphatics, together with the protein content of lymph, that in the gastrointestinal tract reach 4.1g/dl (Ganong 1995). However, it is impressive how the larvae migrate inside the lymph nodes constituted mainly by lymphocytes, being partially retained by a macrophage granulomatous reaction, with or without eosinophils, with rare morphological evidence of larval killing. Only few L3 delayed inside one mesenteric lymph node after 11 days of infection were found degenerated in granulomas. These results suggest that larval hindrance inside the lymph node or lymphatic vessel beyond the physiological limit induces larval death, causing secondary eosinophil infiltration. After spreading all over the arterial system during the final stage of L4/L5 maturation, they selectively choose the mesenteric and mainly the ileocolic and cecal arteries, where they establish the final habitat, causing severe changes in the arterial wall, similar to the atherosclerotic process. Probably the meeting of the male and female is induced by pheromone sex attractants until now showed by about 40 species of nematodes (MacKinnon 1987, Haseeb & Fried 1988). However, it is unknown why they select the ileocolic and cecal arteries, and what is the influence of the gut hormones on the larvogenesis process. In fact, in the human ileum, colon and rectum there is a predominance of somatostatin, glucagon and peptide YY (Polak & Bloom 1992). Basch (1981) successfully cultured the cercarie of Schistosoma mansoni until the pairing stage under medium suplemmented with hormones such as insulin and hydrocortisone. Otherwise, Hata and Kojima (1991) supplemented Waymouth's medium with 10 µg/ml of insulin, 10-8M hydrocortisone, 10-8M β-estradiol, and 30 µg/ml of transferrin, and the addition of these growth factors had no marked effect on A. costaricensis development. The kidney, contrary to the favorable intestinal segments, is rarely affected and when the embolization of the eggs occurs, they present clear signs of degeneration. The A. costaricensis reaches the liver through the branches of the hepatic and/or portal veins, indicating that this organ acts as a crossroad of arterial and venous pathways of the nematode life history. In both situations, L3 grow to full sexual maturity and the eggs are carried into the smallest portal radicals, where, together with L1, they are responsible for the hepatic angiostrongyliasis (a specific paper on this subject is being prepared). Eggs can also reach the gallbladder through the cystic artery that is a branch of the right ramous of hepatic artery. From the intrahepatic portal veins, like S. mansoni, adult worms migrate "upstream" into mesenteric veins (Figs 43, 44) explaining the continuity of egg embolization to intrahepatic portal veins until the end of the experiments (114 days of infection), even in the absence of intrahepatic adult worms. These observations explain the localization of adult worms within a vein of the submucosa of the cecum of a 3-year-old costaricensis boy (Morera et al. 1982). After reviewing the case, the authors also found adult A. costaricensis within the portal veins of the liver. It seems that Angiostrongylus vasorum (Baillet, 1866) completes both its 3rd and 4th molts in the abdominal lymph nodes, and that at least some parasites pass through the liver before reaching the heart and pulmonary arteries (Rosen et al. 1970). According to these authors, this migration appears to occur via the venous system. In our material the adult worms were also in the intrahepatic artery branches from 8th to 29th day of infection. L3 were soon identified inside the intrahepatic portal veins 3 h after the infection. Morera et al. (1982) considered the A. costaricensis adult worm migration to intrahepatic portal veins as a visceral larva migrans-like syndrome. However, our data include this alternative migration to hepatic veins as a normal event of venous portal pathway, where the nematode matures and lays fertile eggs inside the liver. Then, as was stressed in the results, the A. costaricensis adult worms can reach the liver by branches of hepatic artery and portal vein. It is also important to point out the secondary invasion of mesenteric veins by L1, with their subsequent downstream migration to the liver (Figs 31-34). Embolization of L1 to the lungs (not shown on this paper) after the secondary or late invasion of lymphatics in intestinal mucosa and/or in lymph node sinuses by L1 departing from the local larvogenesis originated from embolic eggs might also occur (Figs 27-30). Egg and L1 could also get to the lungs coming from the liver through hepatic veins. The frequent occurrence of embolization to arterial adventitial vessels can explain the prominent periarteritis that evolves to concentric periarterial fibrosis, even in arterial segments deprived of intravascular parasites. This finding also clarifies the occurrence of centripetal (exo - endo) eosinophil arteritis and periflebitis, with the cells moving from the adventitia to the endothelial lining (Graeff-Teixeira et al. 1991). With the exception of Angiostrongylus cantonensis (Chen, 1935), all metastrongylid lungworms that have been studied are believed to reach the lungs entirely via the lymphatic system (Rosen et al. 1970). In fact, using the mouse model, we detected L4 passing through the pulmonary circulation from 3rd to 11th day, being afterwards carried to arteries of various organs by systemic circulation (Mota & Lenzi 1995). The same phenomena were observed also in A. costaricensis infected S. hispidus, however the pre-arterial period was much shorter than in mice because L4 were intra-arterially detected on day 2 (meningeal artery) in one animal, instead of day 5 after the infection as in mice (Mota & Lenzi 1995). This means that the lung passage begins up to 2nd day of infection and may last until the end of the L3 lymphatic phase (11th day). Even Morera (1973) reported L3 arrival in the lungs after 12 and 24 h after infection, drawn through the thoracic duct. A. costaricensis, like the A. vasorum (Rosen et al. 1970), did not develop in the brain of its definitive host as does the closely related parasite, A. cantonensis. The clear detection of adult worms of both sexes inside the left ventricle of the heart, without evidence of pulmonary changes in both cases, induced the conjecture, without any evidence for proof, that they could reach that cavity through pulmonary arteriovenous shunts or persistence/perforation of foramen ovale. Because of that, parasites could migrate to the right cardiac chambers passing through the inferior mesenteric vein branches, rectal plexus or other vein of pelvis, middle hemorrhoidal vein, common iliac vein and inferior vena cava. Another remote possibility could be an intratracheal contamination by the large number of L3 during the oral infection, with further intrapulmonary maturation and invasion of pulmonary vein branches (a study is being performed to verify these hypothesis). All the intracardiac metastrongylid reported in the literature were located in the right chambers of the heart (Rosen et al. 1970). In conclusion, this study confirms the existence of two vascular pathways during the life history of A. costaricensis, as previously showed in the mouse model (Mota & Lenzi 1995) (LYMPHATIC/VENOUS-ARTERIAL and VENOUS PORTAL PATHWAYS), it emphasizes the parasite migration to the liver as an usual event of the cycle, changing the concept of visceral larva migrans-like syndrome as proposed by Morera et al. (1982) and explains most of the parasitic and pathological findings reported sometimes as "odd aspects" (Morera et al. 1982) in humans and naturally infected animals. It also shows that the life history of A. costaricensis is much more complex than the original description (Morera 1973) and still presents some inexplicable events. The study should also stimulate reseachers to investigate more deeply this fascinating nematode which is biochemically and molecularly an almost unknown organism. Figures 51-52 show schematically the vascular distribution of A. costaricensis in the vascular territories and the migratory pathways during the infection. ACKNOWLEDGMENTS To Dr Marcelo Pelajo Machado for his criticism towards the manuscript and for his excellent participation in the theoretical discussion on the subject. To LF Caputo for her technical assistance. REFERENCES
The following images related to this document are available:Photo images[oc05105f39-46.jpg] [oc05105f25-32.jpg] [oc05105f2.jpg] [oc05105f11-16.jpg] [oc05105f52.jpg] [oc05105f17-24.jpg] [oc05105f47-50.jpg] [oc05105f1-4.jpg] [oc05105f1.jpg] [oc05105f3.jpg] [oc05105f5-10.jpg] [oc05105t1.jpg] [oc05105f4.jpg] [oc05105f51.jpg] [oc05105f33-38.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}